Aklilu Bajigo Madalcho, Anna Gazda, Jerzy Szwagrzyk. Ungulate browsing patterns and forage abundance determine the regeneration of palatable tree species: Using new perspectives in assessing ungulate browsing pressure in temperate forests[J]. Forest Ecosystems, 2025, 12(1): 100288. DOI: 10.1016/j.fecs.2024.100288
Citation:
Aklilu Bajigo Madalcho, Anna Gazda, Jerzy Szwagrzyk. Ungulate browsing patterns and forage abundance determine the regeneration of palatable tree species: Using new perspectives in assessing ungulate browsing pressure in temperate forests[J]. Forest Ecosystems, 2025, 12(1): 100288. DOI: 10.1016/j.fecs.2024.100288
Aklilu Bajigo Madalcho, Anna Gazda, Jerzy Szwagrzyk. Ungulate browsing patterns and forage abundance determine the regeneration of palatable tree species: Using new perspectives in assessing ungulate browsing pressure in temperate forests[J]. Forest Ecosystems, 2025, 12(1): 100288. DOI: 10.1016/j.fecs.2024.100288
Citation:
Aklilu Bajigo Madalcho, Anna Gazda, Jerzy Szwagrzyk. Ungulate browsing patterns and forage abundance determine the regeneration of palatable tree species: Using new perspectives in assessing ungulate browsing pressure in temperate forests[J]. Forest Ecosystems, 2025, 12(1): 100288. DOI: 10.1016/j.fecs.2024.100288
Ungulate browsing patterns and forage abundance determine the regeneration of palatable tree species: Using new perspectives in assessing ungulate browsing pressure in temperate forests
Department of Natural Resource Management, College of Agriculture, Wolaita Sodo University, Wolaita Sodo P.O. Box 138, Ethiopia
b.
Department of Forest Biodiversity, Faculty of Forestry, University of Agriculture in Krakow, 31-425, Krakow, Poland
Funds:
Polish National Science Foundation (NCN) (project title: “How do large-scale disturbances influence the relationship between ungulate herbivory and natural regeneration in temperate forests?”) from the Ministry of Science and Higher Education of the Republic of Poland within the framework of statutory activities SUB/040011-D019/2020 of the Department of Forest Biodiversity, University of Agriculture in Krakow2018/31/B/NZ8/02786
Herbivorous ungulates are vital elements of forest ecosystems, yet their impact on tree regeneration and forest dynamics is challenging to assess. Despite their significant influence, few studies measure browsing pressure independently of precise ungulate population data. This study aimed to determine the regeneration mechanisms of palatable tree species in natural forest ecosystems by measuring the forage abundance from young trees of focal species and estimating the browsing pressure exerted by ungulates. We analyzed an extensive dataset of 2,838 tree saplings in two national parks in Poland to examine relationships between variables using a zero-intercept linear regression, and multiple linear regression models, and performed parametric and nonparametric tests. We found that the browsing intensity index (BI) is a good predictor of browsing pressure. Both total abundance and twig dry mass loss due to ungulate browsing were more pronounced in shorter saplings, with these effects decreasing as sapling height increased. Picea abies was the most abundant browse source in the Tatra National Park (TNP), whereas Fagus sylvatica dominated in the Roztocze National Park (RNP). Acer pseudoplatanus and Sorbus aucuparia experienced significantly higher browsing pressure in the TNP, whereas Carpinus betulus was more heavily browsed than other species in the RNP. The increase in browse abundance rapidly reduced ungulate browsing pressure on highly palatable tree species. We conclude that despite the high browsing pressure on the palatable species, the contribution of browse from focal species helps to mitigate this pressure and supports their regeneration in the forest canopy. Therefore, this finding provides valuable insights for managing and conserving tree species vulnerable to ungulate browsing in natural forest ecosystems. In addition, the method we used in this study is novel to estimate browsing pressure and assess the recruitment potential of vulnerable tree species; hence, it can be applied in large-scale studies with similar scenarios.
Browsing of young trees by ungulate herbivores has been recently one of the major issues in forest ecology and management of tree stands (Ammer, 1996; Beguin et al., 2016; Bödeker et al., 2021a, 2021b; Clasen et al., 2015). With the increasing numbers of ungulate herbivores throughout temperate regions of many countries, the problem with the regeneration of palatable tree species increases rapidly (Boulanger et al., 2009; Čermák et al., 2009). In certain conditions, browsing activity can even outweigh the consequences of climate change by altering the dynamics of forest regeneration (Champagne et al., 2021) and inducing profound compositional and structural changes in forests (Didion et al., 2009; Holm et al., 2013).
The problem has been usually addressed by looking at the densities of ungulate herbivores and trying to determine the maximum density that allows for the successful regeneration of trees (Horsley et al., 2003; Tremblay et al., 2007).
However, the browsing pressure exerted by herbivores also depends on the number of young trees available for browsing (Brandner et al., 1990; Mclaren and Peterson, 1994). Browsing interactions on trees can be influenced by the functional nature of forage resources, which can be explained by spatial variations in biomass, structure/height, phenology, and species composition (Fynn et al., 2019). The susceptibility of tree saplings to ungulate browsing can be influenced by the co-existing vegetation, growth rates, and variations in their palatability (Boulanger et al., 2009; Heuze et al., 2005). The composition of young-generation trees with palatable species is more likely to be heavily browsed than less palatable species in the forest ecosystem (Borowski et al., 2021; D'Aprile et al., 2020). While some species are actively avoided by browsing animals, others are selectively targeted (Bodziarczyk et al., 2017). Species like the European hornbeam Carpinus betulus, favored by ungulate browsers, possess the ability to offset losses by generating numerous secondary branches (Samojlik and Kuijper, 2013). Consequently, in forest ecosystems facing notable browsing pressure, the prevalence of Carpinus betulus in the new generation of trees tends to rise and other species exhibit slower development and reduced survival rates (Kuijper et al., 2009). Thus, the effects of ungulate browsing vary significantly based on the availability of young trees and the palatability level (Borkowski, 2004; Brandner et al., 1990; Hidding et al., 2012). Several reports describe the palatability level of different tree species: highly palatable tree species include Sorbus aucuparia, Acer pseudoplatanus, Carpinus betulus, and Abies Alba (Borowski et al., 2021; D'Aprile et al., 2020). Less palatable species include Picea abies (D'Aprile et al., 2020) and Fagus sylvatica (Borowski et al., 2021). According to Angst and Kupferschmid (2023), tree species are classified into five palatability levels, with Level 1 representing less palatable species and Level 5 representing highly palatable species.
Besides, research shows that light conditions influence the relationship between herbivore pressure and the growth or survival of young trees. Trees growing in deep shade struggle to compensate for browsing losses, limiting their potential to develop to higher sizes (Kupferschmid et al., 2015; Mårell et al., 2018; Petritan et al., 2009). However, the high pressure from ungulate herbivores can slow down the height growth of young trees and make them available for repeated browsing even in well-illuminated patches for a long time (Churski et al., 2017).
There are different arguments regarding the relationship between browsing intensity and food availability depending on the type of food and observational scale. According to some research, browsing intensity is reduced by increased browse availability from a higher density of patches (Sample et al., 2023). In contrast, Borowski et al. (2021) hypothesized that patches with abundant forage and species diversity attract ungulates and increase the likelihood of browsing on saplings. However, it is challenging to draw general conclusions as the findings are limited by the methodological approaches used.
Thus far, several methods have been used for studying how ungulate browsing affects the regeneration of trees in forest ecosystems. For example, some studies describe the effects of ungulate browsing on trees by considering ungulate species, density, and foraging behavior (Bohdan et al., 2020; Rhodes and St. Clair, 2018; Sample et al., 2023; Shipley and Spalinger, 1995). Others, however, compute confidence intervals rather than browsing intensity (Bödeker et al., 2021b, Bödeker et al., 2021a), as well as estimate the percentage of edible branches and foliage based on stem base diameter (Bohdan et al., 2020). Furthermore, Akashi et al. (2022) estimated the browsing pressure while accounting for the number of ungulates per area by dividing the number of browsed seedlings per year by the total number of available seedlings.
However, existing methods for assessing the impact of ungulate herbivores on young trees do not provide a complete understanding of the level of damage in natural forest ecosystems. In addition, relying solely on the ungulate population to establish a predator-prey relationship is costly and prone to error due to the open nature of natural forests and the possibility of individual browsers repeatedly visiting the same plant. According to the results of our earlier studies, there are huge differences in the intensity of browsing among tree species (Madalcho et al., 2024), and winter browsing is a continuous process rather than a single event for each sapling (Kupferschmid et al., 2015). Consequently, a customized method is required to close the knowledge gap in determining the browsing pressure that ungulate herbivores exert. Even in situations where the number of ungulates visiting the saplings is unknown, this method aids in understanding how palatable tree species recover under the functional influence of ungulate browsing in natural forest ecosystems.
This study aimed to determine the regeneration mechanisms of palatable tree species in natural forest ecosystems by measuring the forage abundance from young trees of focal species with different palatability levels and estimating the browsing pressure exerted by ungulates.
Our methods involved measuring the total abundance of forage, and the percentage of twig dry mass removed by ungulates for each focal species. Hence, we tested three hypotheses: 1) The relationship between the browsing intensity index (BI) and the percentage loss of twig dry mass to ungulate browsing is significant for focal species. 2) A greater amount of browse availability and higher browsing loss of twig dry mass is at shorter height classes for all focal species. 3) The browsing pressure on the focal palatable tree species decreases with the increase in browse abundance.
2.
Methods
2.1
Site description
Tatra National Park (TNP) and Roztocze National Park (RNP) are characterized by a high density of ungulates. The TNP is located in the Carpathian Mountains and covers an area of approximately 21,000 ha. The bedrock consists of limestone and dolomite in the northern part, and gneiss and granite in the southern. The climate in the lower mountain zone is moderately cool, with an annual temperature of up to 5 ℃ and an annual rainfall of 1,100 mm. The forests of the TNP are dominated by Norway spruce (Picea abies (L.) H.Karst.), European beech (Fagus sylvatica L.) and silver fir (Abies alba Mill.). Among the other species, sycamore maple (Acer pseudoplatanus L.) and rowan (Sorbus aucuparia L.) have been increasing in numbers, especially among the youngest trees. The main forest types are rich beech forests, hygrophilous forests of silver fir and Norway spruce, and forests of silver fir and Norway spruce on acidic substrate (Bodziarczyk et al., 2019; Pielech et al., 2021). The mean density of live trees is 670 individuals per hectare, and the basal area amounts to 27.4 m2·ha−1. The dominant tree species is Norway spruce (88% of the total tree basal area), followed by silver fir (6%) and European beech (3%). Due to the high mortality resulting from windstorms and bark beetle outbreaks, the total amount of coarse woody debris is about 180 m3·ha−1, and 97% of that is Norway spruce. The mortality of canopy trees estimated based on tree volumes amounted to 41% over the last two decades (Bodziarczyk et al., 2019). The guild of ungulate herbivores in the Tatra Mountains consists of chamois (2 ind.·km−2), red deer (1 ind.·km−2), and roe deer (0.5 ind.·km−2) (Tatra National Park archives).
The RNP is located in the central part of the meta-Carpathian upland and covers an area of 8,482 ha, 12% of which is under strict protection. The landscape of the Roztocze Highlands is characterized by long chains of Upper Cretaceous limestone hills reaching 390 m a.s.l. and surrounded by thick layers of postglacial deposits, sand, or loess. The mean yearly temperature is about 8.7 ℃. The yearly amplitude of the mean temperatures often exceeds 22 ℃. The mean annual precipitation is about 700 mm. Out of 40 species native to Poland, 32 naturally occur in the tree stands of the RNP. However, 94% of forest stands consist of three species: scots pine (53% of the total tree volume), silver fir (24%), and European beech (17%). The mean basal area amounted to 37.78 m2·ha−1 (Łukaszewicz et al., 2020). In the RNP the most common herbivores are red deer (5.6 ind.·km−2) and roe deer (4.8 ind.·km−2) (Borowski et al., 2021).
To assess the amount of browse available from young trees for ungulate herbivores, and the browsing pressure, first we measured young trees in the lab to develop equations that describe the allometric relationships between tree height, stem thickness, and other morphological traits with the dry mass of twigs available for browsing (Madalcho et al., 2024). Second, we set up research plots in the forest as belt transects to measure sapling parameters. Then, we calculated the dry mass of available twigs, the amount of twig dry mass loss to ungulate browsing, and the percentage of twig dry mass loss to ungulate browsing for each tree species by using these measurements and allometric relationships. In this work, a “twig dry mass (g)” stands for a browse abundance, and the “percentage of twig dry mass loss to ungulate browsing (%)” stands for browsing pressure. We classified our focal tree species into three functional groups based on palatability (Angst and Kupferschmid, 2023) to examine the relationship between browse abundance and ungulate browsing pressure. Sorbus aucuparia, Acer pseudoplatanus, Carpinus betulus, and Abies Alba were classified as highly palatable, Fagus sylvatica as moderately palatable, and Picea abies as less palatable tree species.
2.2
Field data collection
At each study site, we set up 22 belt transects. The transects were 30 m long and 5 m wide. They were established using a stratified random approach in areas characterized by extensive tree regeneration, at least 100 m distant from roads and hiking trails, in strictly protected areas or the zone of active management, but not affected by human activities in recent years. Most of them run from the gap to the dense forest (with the forest/gap edge in the middle of the transect).
The forests contain diverse winter food sources like raspberry, blackberry, and blueberry. However, our research focuses on saplings of economically important species in our study areas. We recorded 15 tree species, with six - Picea abies, Fagus sylvatica, Abies alba, Acer pseudoplatanus, Sorbus aucuparia, and Carpinus betulus - present in sufficient numbers for meaningful statistical analysis. A maximum of 30 saplings (individuals from 0.5 to 3.0 m in height) of each tree species present at the study sites were sampled in each transect. To do that, we put a measuring tape along the long axis in the middle of the transect and then searched for saplings that were closest to the line, measuring distances from the transect axis to the stem base of individual saplings. For less abundant species we measured saplings over the entire transect area and in many cases the number of measured saplings was still less than 30 individuals although all individuals occurring within the transect were measured.
In each sapling, we measured its height, diameter at the stem base, the diameters of all shoots at the point where they were browsed, and the height of this point. We measured only shoots that were fully lignified to avoid the tiny shoots that could have been eaten either by rodents or by birds. In each case, we started recording the browsed shoots from the top of the tree saplings and measured at most 50 browsed shoots per transect for each species. We measured only the new browsing, as the signs of old browsing in some species are hard to identify. Fieldwork was carried out in late spring-early summer of 2020.
2.3
Data processing
We estimated twig dry mass for the saplings using species-specific allometric equations that relate dry mass to height class categories (Madalcho et al., 2023). This approach allowed us to calculate the potential twig dry mass for each tree species at the study sites. The calculation included all saplings of the target tree species, both browsed and unbrowsed, that fell within the height class range defined in our study to determine the total potential twig dry mass per unit area.
The twig dry mass removal due to ungulate browsing was determined using allometric equations based on twig diameter classes (Madalcho et al., 2023). The branches of tree saplings with thicknesses between 2 and 7 mm were classified into five diameter classes: 2, 3, 4, 5, and 6 mm. The vertical distribution of twig dry mass consumed by ungulates was analyzed by grouping measurements into three height classes: ≤1, 1–2, and 2–3 m. These height classes corresponded to the positions where the browsed shoot diameters were measured. The vertical distribution pattern of ungulate browsing was assessed by considering both the diameter of the browsed twigs and the height at which these diameters were measured.
We calculated the percent twig dry mass loss of each species by dividing the actual twig dry mass loss (g) by the total amount of twig dry mass (g) per transect, then multiplying the result by 100. This metric reflects the actual browsing pressure, whereas the BI measures theoretical browsing pressure on each species.
Browsing pressure =( Twig dry mass lost to ungulate browsing (g) Browse abundance (g))×100
(1)
BI=N∑i=1d2iD2
(2)
where d is the diameter of the browsed shoot (mm), N is the number of browsed shoots per plant, and D (mm) is the basal diameter of the tree sapling. As a result, the mean BI values of individual saplings per transect were considered to represent the species-level BI, which was then utilized to assess its relationship with the proportion of twig dry mass removed by ungulates for each species.
The twig dry mass that remained after ungulate browsing was calculated as the difference between the potential total twig dry mass and the twig dry mass consumed by ungulates for each species.
2.4
Data analysis
The relationship between browse availability and ungulate browsing was analyzed using a linear regression model with a zero intercept, reflecting the logical assumption that no browsing can occur without available browse material. The formula y = ax is expressed as y ~ 0 + ax. The vertical distribution of twig dry mass (g), the amount of twig dry mass removed by ungulate browsing (g), and the percentage of twig dry mass loss to ungulate browsing (hereafter, browsing pressure) were analyzed using one-way ANOVA, and Bonferroni tests of post-hoc were computed.
We considered a functional group-level analysis to observe the interactive effect of total browse abundance and palatability on the ungulate browsing pressure on the focal tree species. Hence, a multiple linear regression model with an interaction term was used to analyze the effect of browse abundance, species palatability, and their interaction term on ungulate browsing pressure on tree species. This model is often referred to as a two-way interaction model or moderated multiple regression.
Multiple linear regression
Y=β0+β1X1+β2X2+β3(X1X2)+ε
(3)
where Y is the dependent variable (ungulate browsing pressure), X1 is the first independent variable (browse abundance), X2 is the second independent variable (species palatability), X1X2 is the interaction term, β0 is the intercept, β1, β2, and β3 are the regression coefficients, and ε is the error term.
We compared browsing pressure and remaining twig dry mass after ungulate browsing (g) between the study sites for the tree species using a nonparametric test (Mann-Whitney U). Hence, the group with a higher median rank was regarded as having a greater value. The difference between tree species for these variables in each study area was calculated using one-way ANOVA, and post-hoc mean separation was computed with the Scheffé test. The data was analyzed and visualized using R-version 4.3.1, Statistica (TIBCO-version 14.0.0.15), and Microsoft Excel.
3.
Results
3.1
Distribution of total browse abundance at each study site
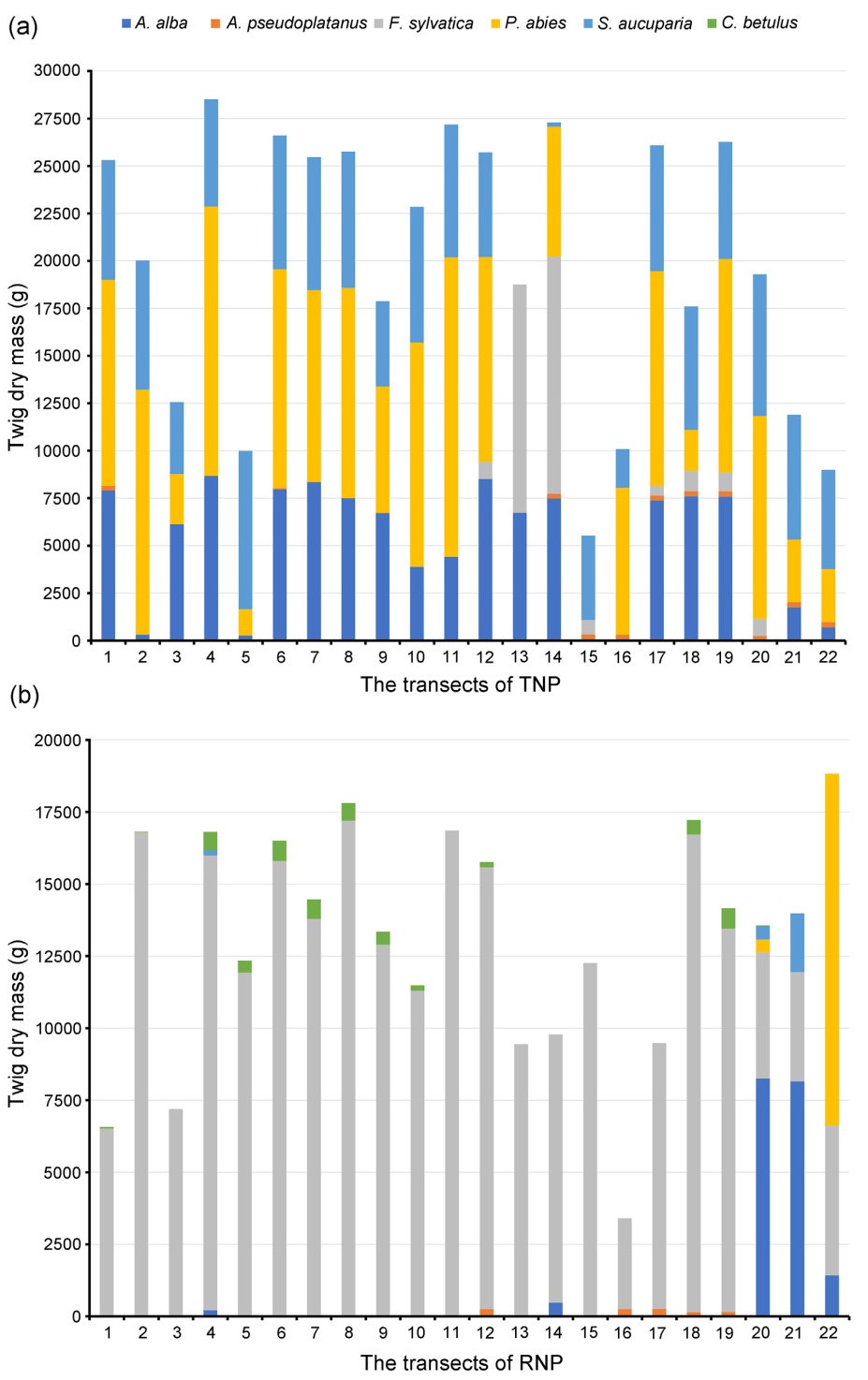
Fig. 1a and b presents the distribution of browse abundance at each study site and within each transect. For each species, twig dry mass per transect represents the sum of the dry mass of up to 30 individual tree saplings. The distribution of twig dry mass exhibited notable differences between the tree species and across the study sites (Fig. 1).
Figure
1.
The distribution of twig dry mass abundance per transect for each tree species in the Tatra National Park (a) and the Roztocze National Park (b). This general overview of transect-level distribution data helps to interpret the species-level analysis.
3.2
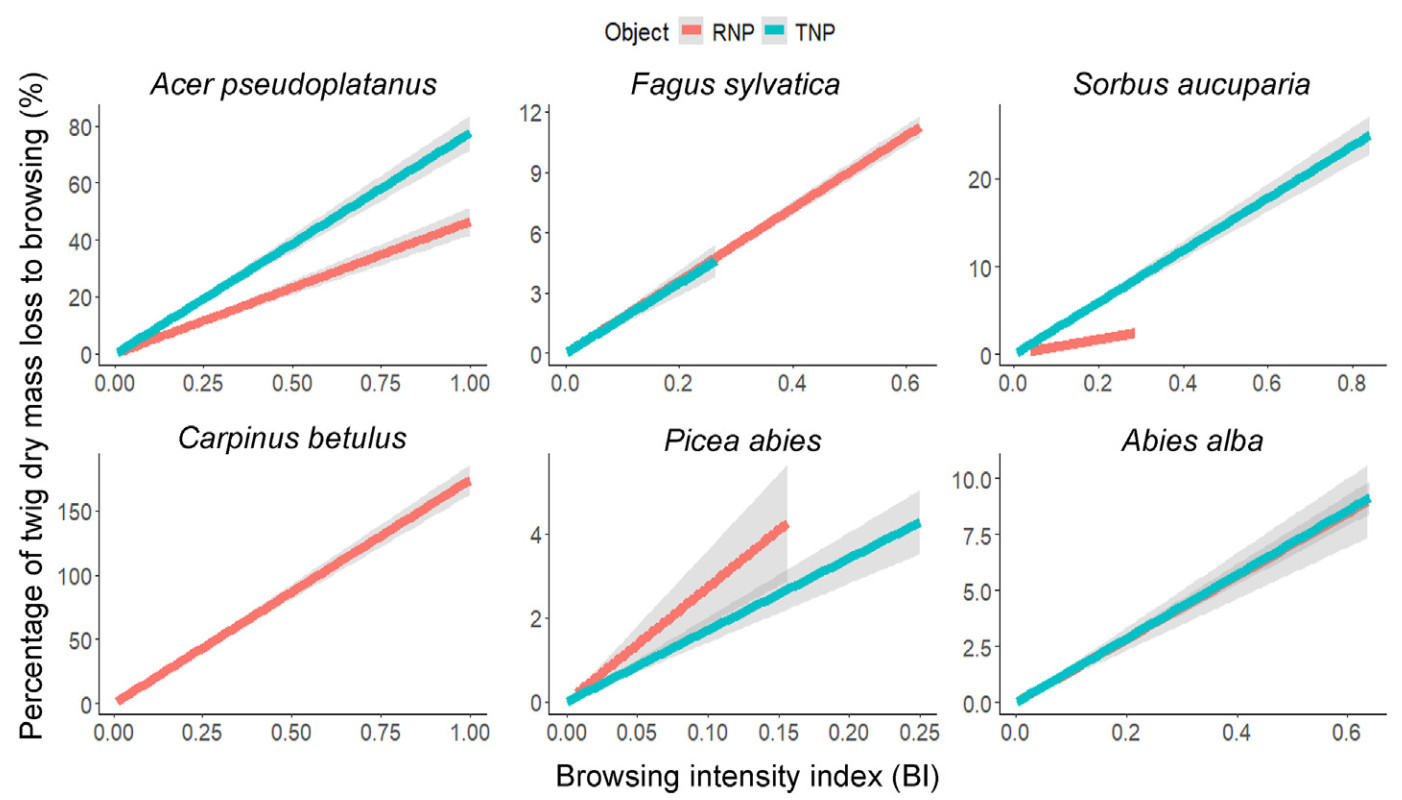
The relationship between the browsing intensity index (BI) and the browsing pressure
The relationship between BI and the ungulate browsing pressure was strongly significant for all species (Table 1). However, the strength of the relationships between these variables was significantly varied only for Acer pseudoplatanus and Sorbus aucuparia in the study areas. As a result, a stronger relationship was observed in the TNP than the RNP for these species, whereas there was no significant variation among the study sites for the rest of the focal species (Fig. 2).
Table
1.
A zero-intercept linear model for the association between browsing intensity index (BI) and the percentage of twig dry mass of a sapling eliminated by ungulate browsing. BI as a predictor variable was used to estimate the ungulate browsing pressure at each of the study sites.
3.3
Vertical distribution of twig dry mass abundance and ungulate browsing loss
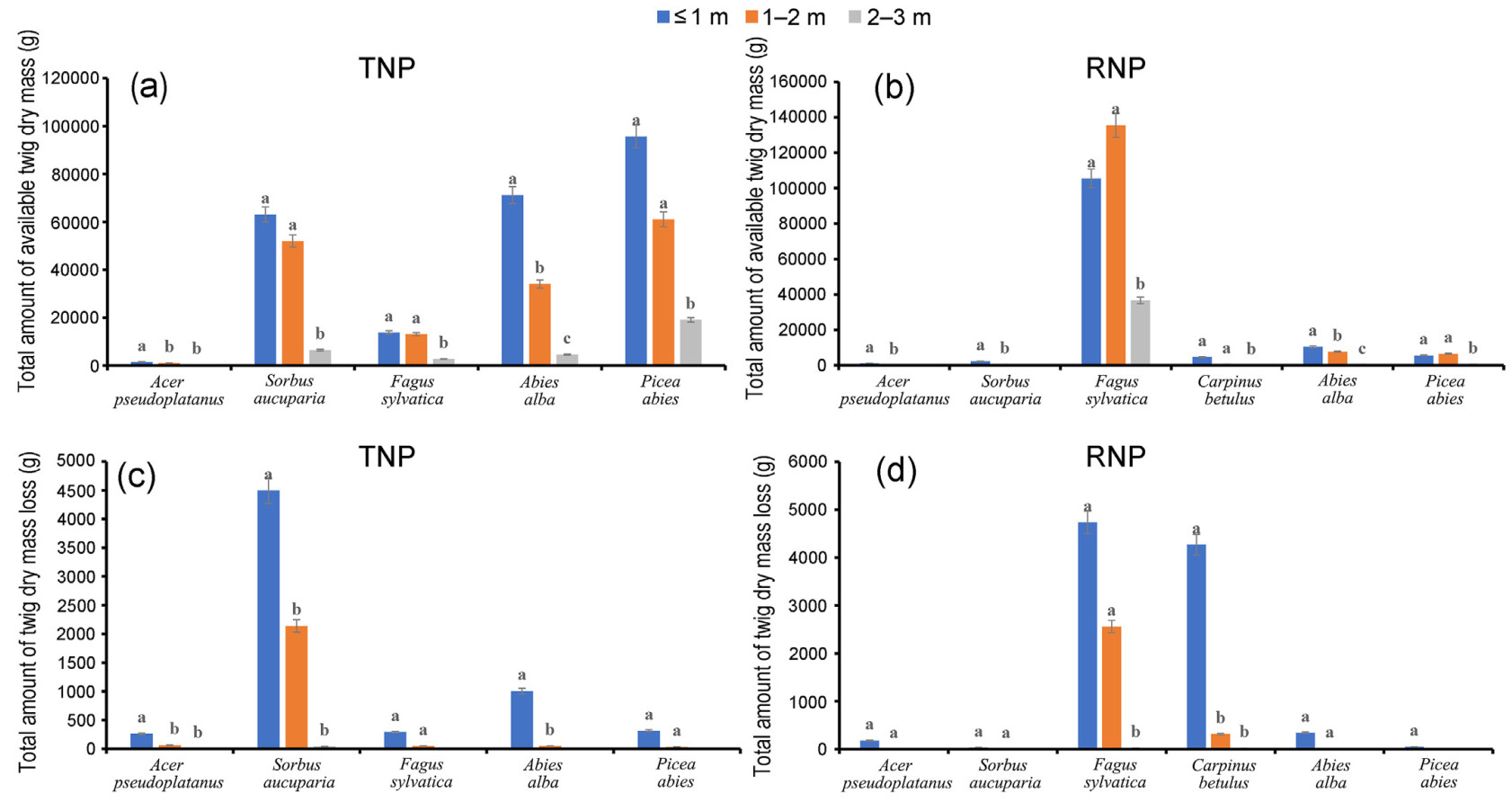
The total abundance of twig dry mass decreased along the height classes for all species in both TNP and RNP, as short saplings were much more numerous than taller ones. In the TNP, the twig dry mass was distributed over all height classes for all tree species. For Acer pseudoplatanus and Abies alba, a significantly higher amount of browse was observed in the sapling height of ≤1 m, while there was no significant difference between the taller height classes. Also for Sorbus aucuparia, Fagus sylvatica, and Picea abies, the twig dry mass was significantly higher at the sapling height of ≤2 m. However, there were no significant differences between the height classes below 1 and 1–2 m (Fig. 3a). In the RNP, twig dry mass was distributed in all height classes for all species except Acer pseudoplatanus and Sorbus aucuparia, which were not found in heights >2 m. For Acer pseudoplatanus, Sorbus aucuparia, and Abies alba, twig dry mass was significantly higher at heights ≤1 m and decreased with height. In the height classes below 2 m, the twig dry mass (g) was significantly higher for Fagus sylvatica, Carpinus betulus, and P. abies, while it did not differ significantly between the height classes ≤1 and 1–2 m (Fig. 3b).
Figure
3.
The vertical distribution of total twig dry mass availability (a and b) and the twig dry mass loss by ungulate herbivores (c and d) is categorized into three height classes: ≤1, 1–2, and 2–3 m. The height classes for twig dry mass loss by ungulate herbivores are based on the height at which the twig was browsed. The maximum height of the saplings is 3 m. According to the results of the post-hoc Scheffé test (p < 0.05), different letters indicate significant differences in values within height classes.
The vertical distribution of twig dry mass loss (g) due to ungulate browsing varied by tree species and study area. In the TNP, Acer pseudoplatanus, Sorbus aucuparia, and Abies alba showed significantly higher twig dry mass loss (g) at heights ≤1 m. In addition, Acer pseudoplatanus and Sorbus aucuparia experienced browsing damage at taller heights of up to 3 m at this site. On the other hand, for Fagus sylvatica and Picea abies, there was no significant difference in browsing loss between the shorter height classes, and twig dry mass loss did not extend to 3 m twig height (Fig. 3c). In the RNP, Acer pseudoplatanus, Sorbus aucuparia, and Abies alba showed browsing loss up to heights of ≤2 m with no significant difference between these shorter height classes, while Picea abies had damage limited to ≤1 m. In contrast, Fagus sylvatica and Carpinus betulus had significantly higher browsing loss to twig dry mass at ≤1 m sapling browsing height, which decreased with height up to 3 m (Fig. 3d).
3.4
Browse abundance and browsing pressure on the focal tree species across forest communities
3.4.1
The effect of browse abundance on ungulate browsing pressure for palatable species
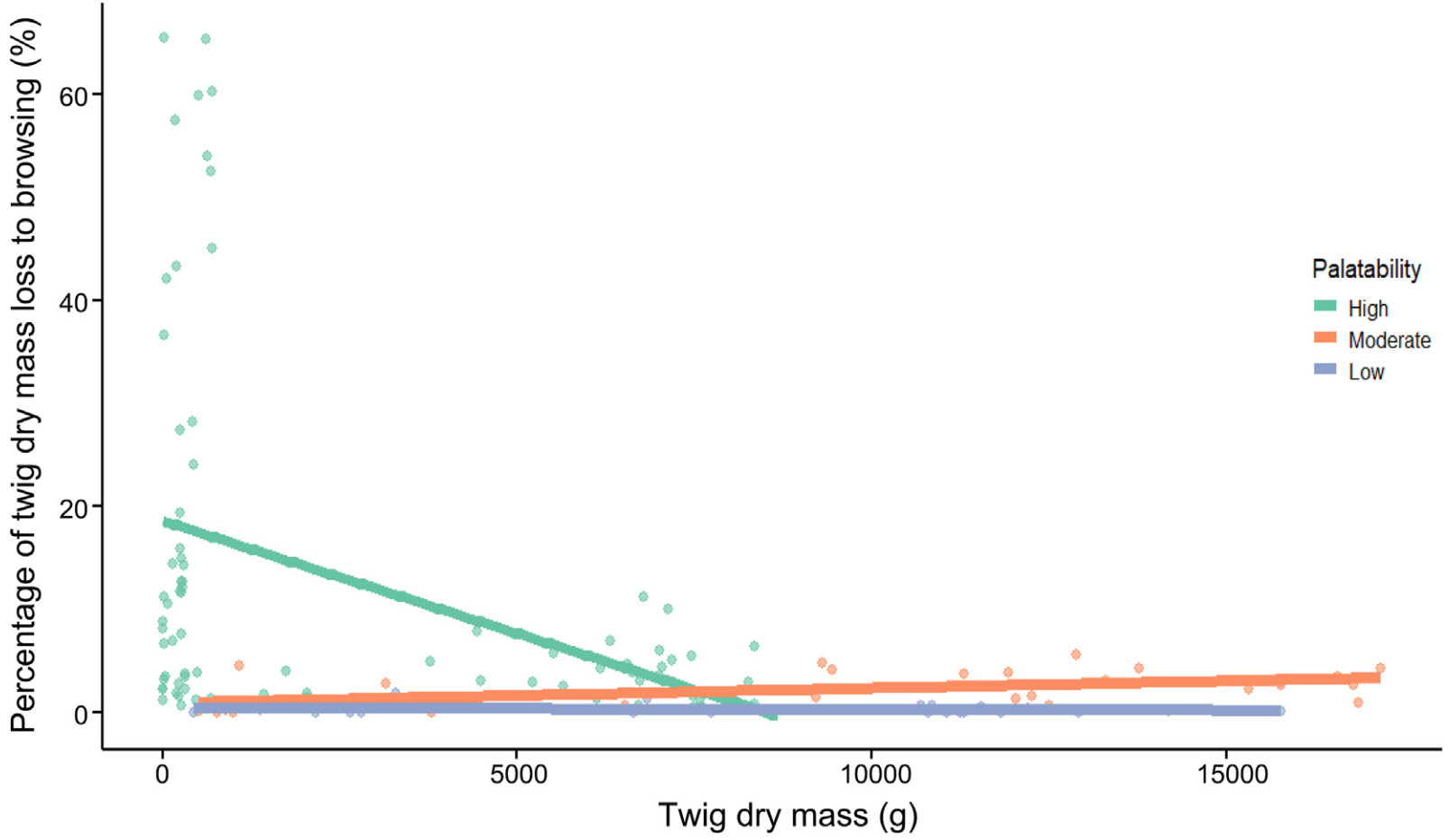
The browsing pressure on highly palatable species decreased rapidly with increasing browse abundance, while pressure on moderately and less palatable species increased gradually as browse abundance increased (Fig. 4).
Figure
4.
The effect of twig dry mass (the browse abundance) and the species palatability on the percentage of twig dry mass loss to ungulate browsing (browsing pressure) of focal tree saplings.
The model examined the relationship between browsing pressure and two predictors: browse abundance and palatability, with their interaction being significant. The intercept (18.63) represents the expected browsing pressure when browse abundance increases by zero units and palatability is at its reference level (presumably “high”). For each unit increase in browse abundance, browsing pressure decreased by 0.0022 units for highly palatable species. This effect is statistically significant (p < 0.001). On the other hand, low palatability reduced browsing pressure by 18.20 units compared to the reference level (p < 0.01) and moderate palatability reduced browsing loss by 17.70 units compared to the reference level (p < 0.001).
The interaction terms showed that the effect of browse abundance on browsing pressure varied across palatability levels. For species with low palatability (p < 0.01) and those with moderate palatability (p < 0.001), the effect of browse abundance on browsing pressure was significantly different from that of highly palatable species (Table 2).
Table
2.
The effect of browse abundance and species palatability level on browsing pressure of tree species. The coefficients for “low-palatability” and “moderate-palatability” show how these levels differ from the high palatability level.
3.4.2
Differences in the total browse abundance between the study sites and among tree species
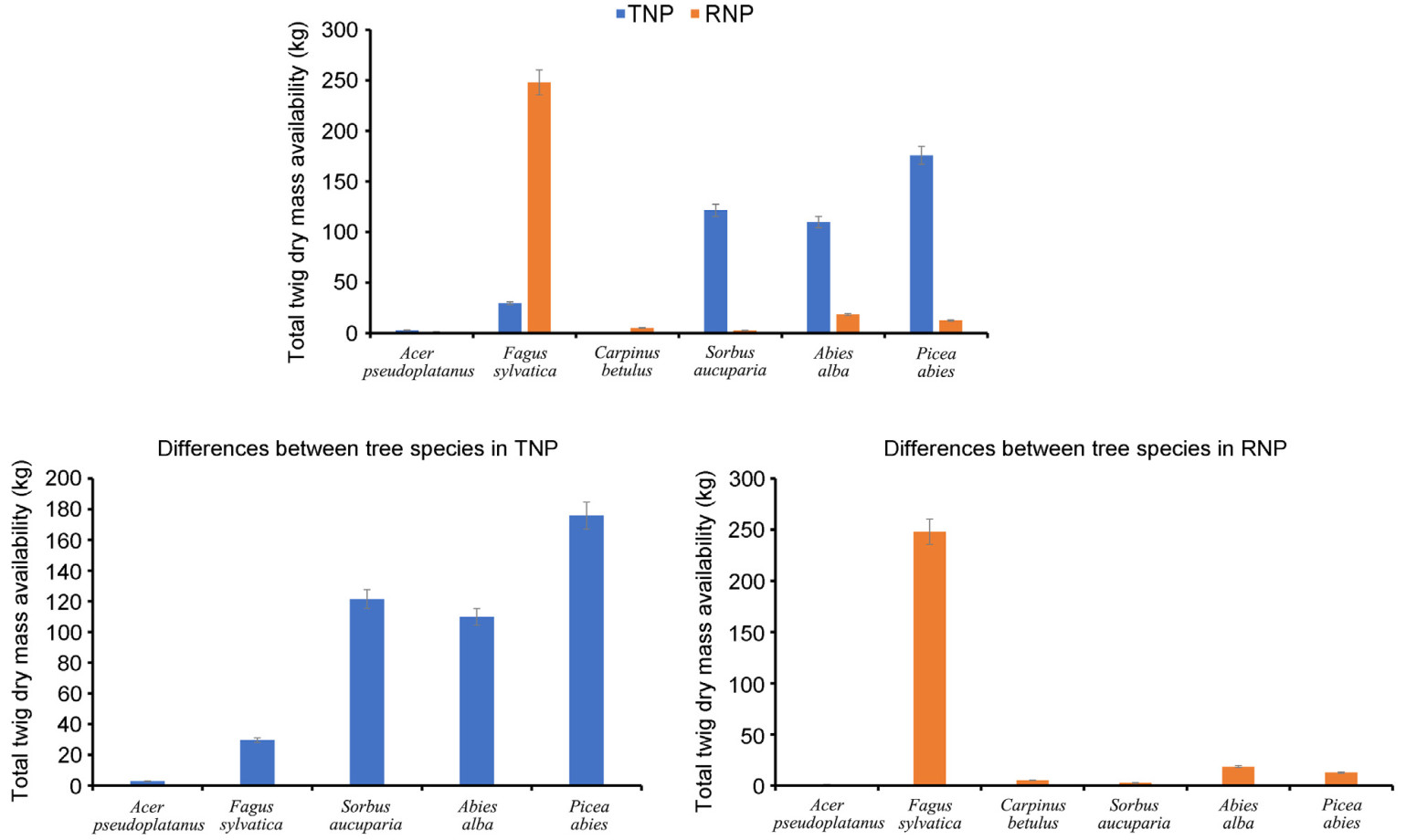
There was a large difference in total browse abundance, measured as twig dry mass (kg) per study site, between the two National Parks. In TNP, the abundance of Picea abies, Abies alba, Sorbus aucuparia, and Acer pseudoplatanus was much higher than in RNP, where Fagus sylvatica was much more abundant. Within the TNP, Picea abies had the highest total browse abundance, followed by Sorbus aucuparia and Abies alba. In contrast, Acer pseudoplatanus and Fagus sylvatica had very low browse abundance in this park. On the other hand, Fagus sylvatica had the highest total browse abundance and spatial distribution in the RNP, followed by Abies alba (Fig. 5).
Figure
5.
Variations in the total browse abundance across the study sites and among tree species on each site (3,300 m2).
3.4.3
Differences in the ungulate browsing pressure between the study sites and among the tree species
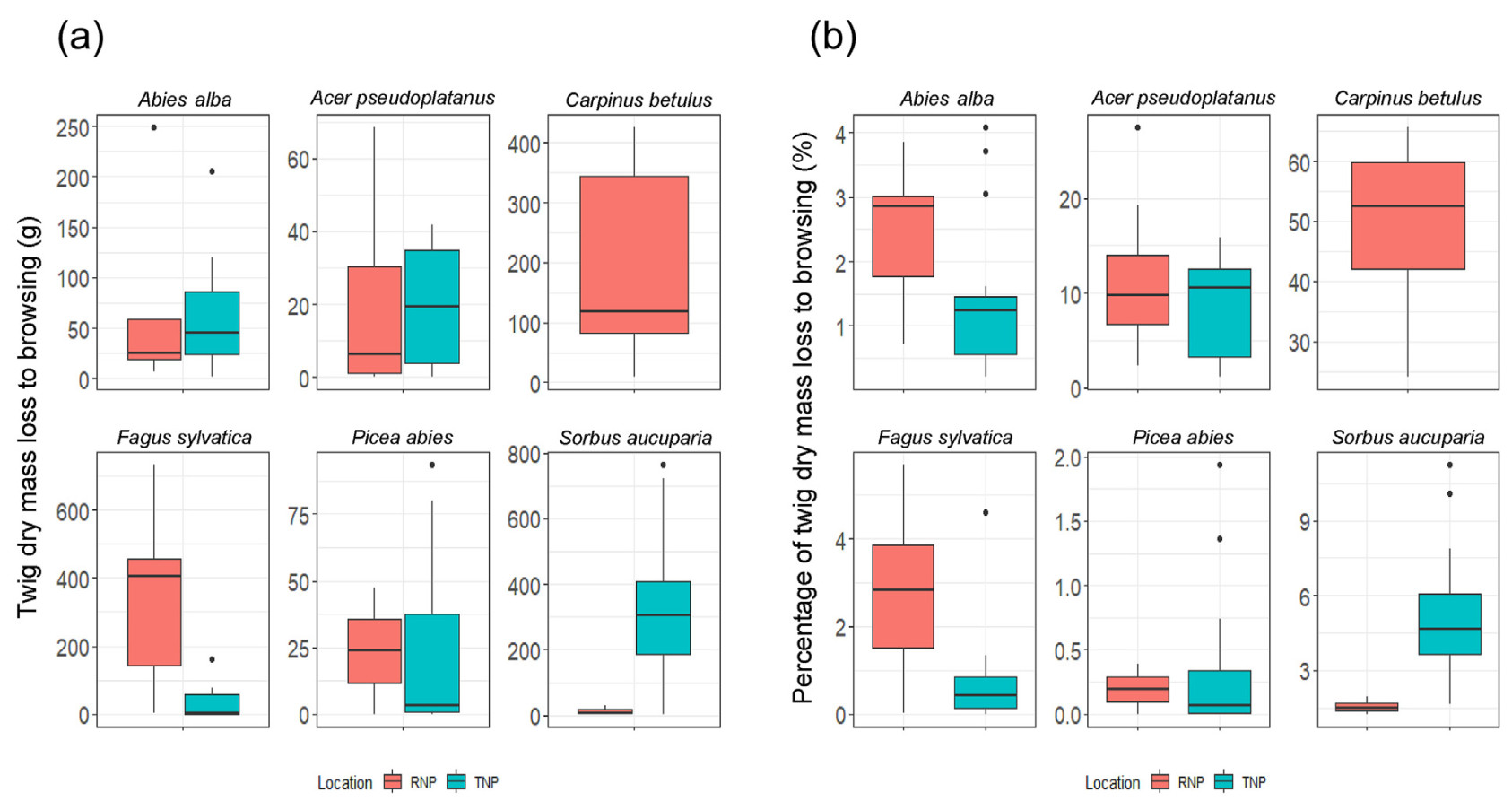
The actual loss of twig dry mass (Fig. 6a) and the percentage loss of twig dry mass due to ungulate browsing (Fig. 6b) were significantly different across the study sites for Fagus sylvatica and Sorbus aucuparia. Fagus sylvatica experienced considerably more browsing loss of twig dry mass in the RNP compared to the TNP. Conversely, Sorbus aucuparia experienced significantly more damage in the TNP. The pattern of browsing pressure mirrored the browsing damage observed in both species. There was no significant difference between the study sites for the other tree species (Table 3).
Figure
6.
(a) The actual loss of twig dry mass, and (b) the browsing pressure to ungulate herbivores: Comparison between the two study locations.
Table
3.
Wilcoxon rank sum test with continuity correction was used to compare twig dry mass loss due to ungulate browsing and the browsing pressure between RNP and TNP. A Non-parametric test (Mann Whitney U) (p < 0.05).
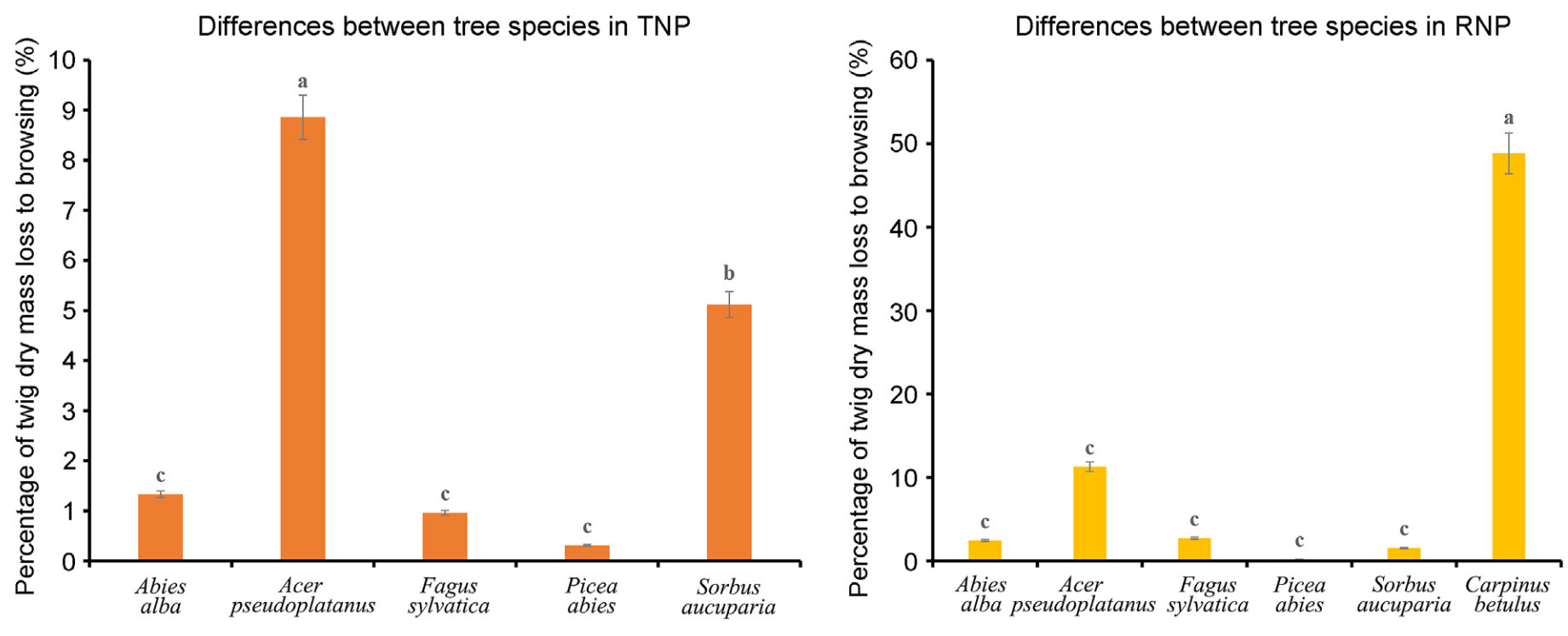
The browsing pressure exhibited variation among tree species across study sites. In the TNP, Acer pseudoplatanus was found to experience the highest level of browsing pressure, followed by Sorbus aucuparia. The browsing pressure on Fagus sylvatica, Abies alba, and Picea abies was similar and relatively low, with no significant differences between the three species. At the RNP site, Carpinus betulus exhibited notably elevated levels of browsing pressure, while all other tree species demonstrated comparably lower browsing pressure (Fig. 7).
Figure
7.
Comparison of tree species concerning ungulate browsing pressure at each study site. According to Scheffé's post-hoc test (p < 0.05), different letters indicate significant differences in browsing pressure among tree species within each study site.
3.5
The remaining twig dry mass after ungulate browsing in different sites and species
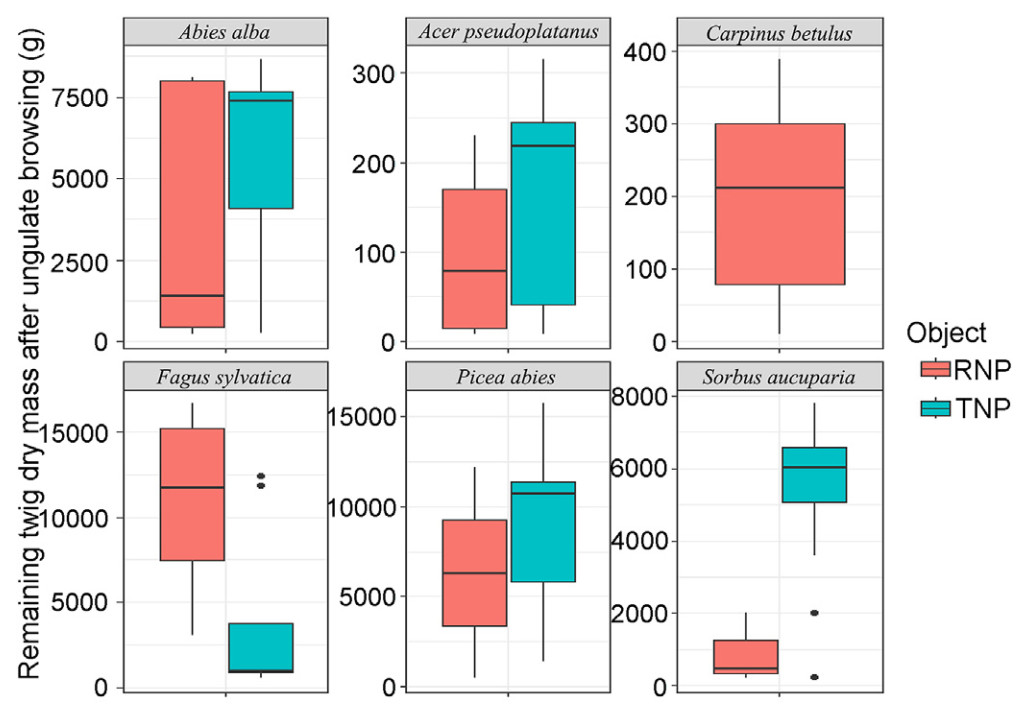
Wilcoxon rank sum test results indicated a significant difference in the amount of twig dry mass remaining after ungulate browsing between TNP and RNP for Fagus sylvatica and Sorbus aucuparia (Table 4). Specifically, Fagus sylvatica had significantly more twig dry mass remaining in RNP, while Sorbus aucuparia had significantly more in TNP (Fig. 8). However, no significant differences between the study sites were observed for Acer pseudoplatanus and conifers.
Table
4.
Wilcoxon rank sum test with continuity correction for comparison between twig dry mass remaining after ungulate browsing in RNP and TNP. A nonparametric test (Mann Whitney U) with 95% confidence.
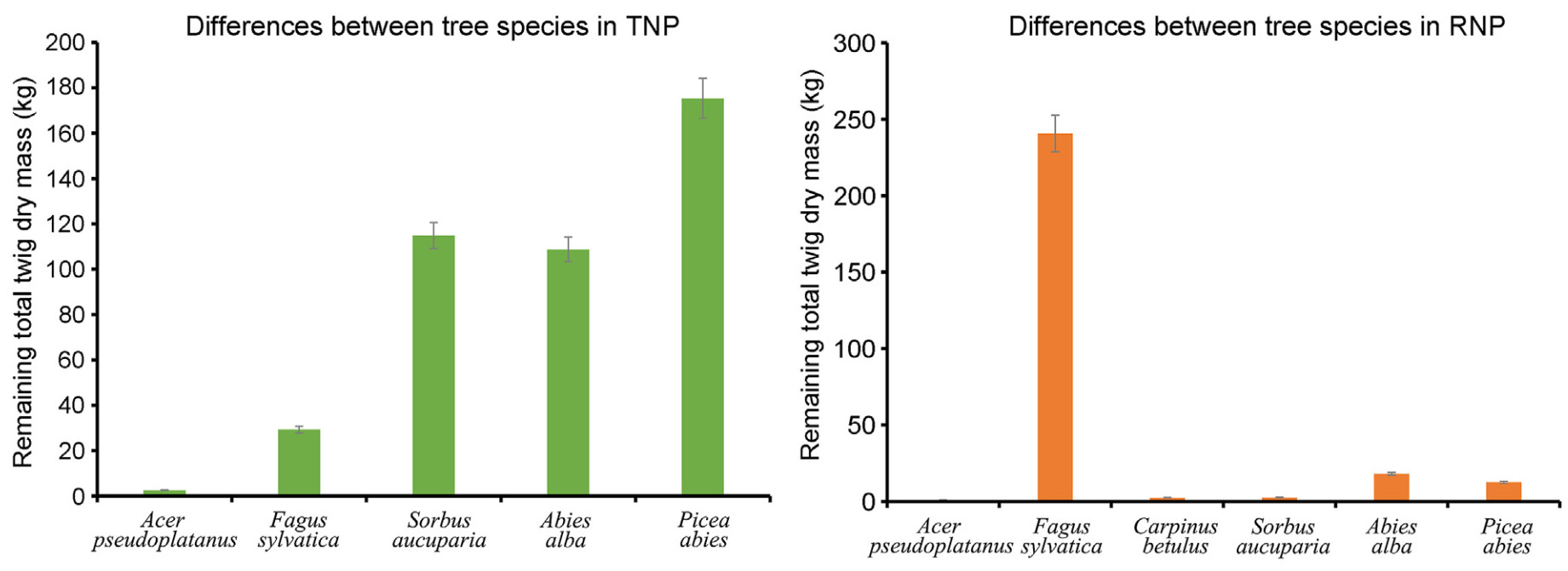
The comparison among the species showed that Picea abies had a much higher dry mass of twigs left after ungulate browsing followed by Sorbus aucuparia and Abies alba in the TNP, while Acer pseudoplatanus had a significantly lowest twig dry mass. Whereas in the RNP, the dry mass of the remaining twigs of Fagus sylvatica was significantly higher than that of the other species (Fig. 9).
Figure
9.
Comparison of tree species according to the dry mass of twigs left over after ungulate browsing in each of the study sites (3,300 m2).
4.1
The relationship between the browsing intensity index (BI) and the browsing pressure
The browsing intensity index (BI) is a theoretical measure of ungulate browsing pressure on tree saplings. However, the browsing pressure we calculated, reflects the percentage loss of twig dry mass, which directly quantifies the actual loss in twig dry mass from what is available for browsing. In terms of practical application, BI is easier to measure in the field, whereas calculating browsing pressure involves a lengthy process.
Therefore, the strong positive correlation between BI and ungulate browsing pressure on the tree species suggests that BI stands out as an alternative and simpler method for estimating the actual browsing pressure on tree saplings. The model relationship between these variables is also relevant for large-scale studies of browsing pressure on the tree species. Connecting these variables is necessary to propose a simpler method and to assess the actual browsing pressure, as the proportion of shoots consumed by ungulates offers reliable estimates of food availability and ungulate food consumption (Gaudry et al., 2022). Furthermore, the model is a better fit for Acer pseudoplatanus and Sorbus aucuparia in the TNP, however, for the rest of the focal species, the quality of capturing the error does not vary across the study sites.
4.2
Vertical distribution of browse abundance and ungulate browsing damage
Browse abundance was greater in shorter saplings (height classes ≤1 and 1–2 m) for all focal tree species. This may be due to the higher density of shorter height classes in both areas. The higher density of shorter saplings often results from their increased susceptibility to browsing. This density may lead to competition for light, water, and nutrients, and slow the height growth (Bödeker et al., 2023; Nopp-Mayr et al., 2020). The current results support our hypothesis that browsing slows sapling growth and maintains them at shorter heights for longer periods.
In the TNP, browse abundance was distributed across all height classes up to 3 m for each species, but it declined significantly as height increased. This pattern may be due to a historical natural disturbance that created a canopy gap, encouraging the growth of light-demanding broadleaf species. On the other hand, light-demanding species such as Acer pseudoplatanus and Sorbus aucuparia do not have browse availability above 2 m in the RNP, where natural disturbances are minimal (Fig. 3a and b). Light availability is critical for sapling growth. Light-demanding species can thrive in canopy gaps created by disturbance, but if these gaps close and browsing pressure remains high, their growth may be restricted (Knapp et al., 2019).
Our results show that the vertical distribution of twig dry mass loss due to ungulate browsing is high on the saplings of all species at shorter heights. However, there is variation in the vertical distribution of browsing loss of twig dry mass among the tree species and between the study sites. This variation is likely influenced by multiple interacting biophysical factors such as elevation, competing sources of browse resources, and the wild ungulate population (Bernard et al., 2024; Hardalau et al., 2024; Pranzini et al., 2024).
We find a consistent pattern between the browse abundance of focal tree species and the vertical distribution of twig dry mass loss from ungulate browsing. Thus, the significantly higher twig dry mass loss from browsing at heights of 1 m or less is observed in Acer pseudoplatanus, Sorbus aucuparia, and Abies alba in the case of TNP. This finding supports the earlier research, including D'Aprile et al. (2020), which discovered that saplings under 150 cm tall are primarily impacted by ungulate browsing. Similarly, the most heavily browsed group of saplings are shorter ones, according to Szwagrzyk et al. (2020).
Ungulate herbivores, such as deer, play a significant role in shaping forest structure by affecting tree regeneration and growth. Browsing tends to be more intense on shorter saplings because they are within reach of herbivores. This results in slower growth rates and can keep saplings at shorter heights for longer periods, which could increase tree mortality (Bödeker et al., 2023).
However, the results also suggest that the palatability of the species may influence the vertical distribution of dry mass loss of twigs due to browsing by ungulates. For example, for palatable species such as Acer pseudoplatanus and Sorbus aucuparia, the vertical distribution of twig dry mass loss due to ungulate browsing on tree species reaches heights of up to 3 m. On the other hand, less palatable species such as Picea abies, Abies alba, and Fagus sylvatica do not show browsing incidence at heights above 2 m in the TNP.
In the RNP, browsing is restricted to heights of up to 2 m for Acer pseudoplatanus and Sorbus aucuparia, resulting in no browsing damage above this height. Nevertheless, the browsing loss of twig dry mass is comparable at height classes ≤1 and 1–2 m, suggesting that both species are subject to comparable browsing pressure at these heights. The lack of twig dry mass at a height taller than 2 m for these palatable species may be primarily associated with the influence of high browsing pressure in the past (Kupferschmid et al., 2019; Szwagrzyk et al., 2020), and low light intensity under the forest canopy (De Pauw et al., 2022). In contrast, Abies alba and Picea abies, which are shade-tolerant, have browse available in all height classes. Rather, ungulate browsers tend to avoid the taller saplings of these species. Fagus sylvatica and Carpinus betulus, however, experience more vertically distributed browse loss due to ungulate herbivores in this forest ecosystem. They suffer significantly greater twig dry mass loss at heights up to 1 m, with a notable decline in loss as the height increases.
In general, the results of vertical distribution of twig dry mass loss to ungulate browsing on tree species indicate that, despite inter-species variation, saplings of palatable species show increased resilience to browsing intensity due to the burden shared by less palatable species. These species contribute by increasing the abundance of alternative browse, and reducing damage to the saplings of palatable species, as observed in the case of TNP (Fig. 3c). Tree species composition and browse availability significantly influence browsing intensity on individual species (Faison et al., 2016; Ohse et al., 2017).
4.3
The relationship between browse abundance and ungulate browsing pressure on the focal tree species across forest communities
4.3.1
The effect of browse abundance on the ungulate browsing pressure on palatable species
The availability of browse resources influences the browsing behavior of ungulates on each tree species. Consequently, as browse abundance for a particular species increases, ungulates tend to exert greater browsing pressure on that species, consistent with the findings of Gaudry et al. (2022).
However, an increase in browse abundance at the community level leads to a decrease in browsing pressure on more palatable species. Highly palatable species experience the greatest baseline browsing loss, but as browse abundance increases, their browsing pressure decreases more rapidly than less palatable species, showing a strong negative relationship between browse abundance and browsing pressure; and it confirms our third hypothesis. This observation supports the argument that browsing pressure decreases as the overall abundance of browse resources increases (Sample et al., 2023). Our result shows that as the total browse abundance of focal species increases, there is a marked decrease in browsing pressure on more palatable species. This effect is particularly pronounced for highly palatable species such as Acer pseudoplatanus, Sorbus aucuparia, Abies alba, and Carpinus betulus, which experience a rapid decline in browsing pressure as total browse abundance increases. In contrast, medium palatable species such as Fagus sylvatica show a more gradual decline in browsing pressure than less palatable species such as Picea abies.
This interaction suggests that an abundance of browse resources plays a critical role in modulating ungulate foraging behavior. When browse is abundant, ungulates distribute their foraging more evenly across available species, potentially reducing the intense pressure on highly palatable species. Conversely, when browse is scarce, highly palatable species bear the brunt of browsing pressure, likely due to their preferred nutritional qualities.
4.3.2
Comparison of browse abundance across study sites and among tree species
The comparison of browse abundance between the two national parks shows differences for each tree species. In particular, the higher browse abundance of Picea abies, Sorbus aucuparia, Abies alba, and Acer pseudoplatanus in the TNP compared to the RNP can be attributed to differences in forest gaps. In the TNP, most of the transects (95.5%) contain Sorbus aucuparia, which thrives in large canopy gaps and open areas, although they are also able to survive under forest canopy, but in stunted form (Hamberg et al., 2015; Petritan et al., 2009; Zywiec et al., 2013; Żywiec and Ledwoń, 2008). The TNP has much larger gaps due to the historic large-scale natural disturbances that have occurred since 2004 on the spruce-dominated forest stand (Pajtík et al., 2015; Sproull et al., 2015). On the other hand, the saplings of Fagus sylvatica are distributed in all transects in the RNP (Fig. 1). This shade-tolerant and highly competitive species is also less browsed than the other deciduous tree species and dominates the young generation of trees. In TNP it is less abundant, due to a combination of environmental and historical factors (Pielech et al., 2021). The lower browse abundance of Sorbus aucuparia in the RNP may be associated with a high browsing pressure due to the higher herbivore density in the area. Chronic impacts from ungulate browsing can reduce the abundance of palatable tree species by reducing the population density and structural development (Didion et al., 2009). When comparing tree species in the TNP, Picea abies, followed by Sorbus aucuparia and Abies alba, provides a substantial amount of browse, while Acer pseudoplatanus contributes the least.
4.3.3
Comparison of ungulate browsing pressure across study sites and between tree species
The ungulate browsing pressure varies across the study sites in the case of Fagus sylvatica and Sorbus aucuparia. The higher browsing pressure on Fagus sylvatica in the RNP may be related to the differences in the distribution of browse and species composition between the study sites following the differences in the canopy gaps. Despite the negative relationship between browse abundance and browsing pressure, the shade-tolerant Fagus sylvatica saplings dominate most of the transects in the study area in the RNP and remain the main browse source for ungulate herbivores. Similarly, the highly palatable Sorbus aucuparia experienced more browsing pressure in the TNP than in the RNP, which may be because the distribution of the species was limited to a few transects in the RNP. Different findings show that browsing pressure on tree species in a given site is influenced by the interacting factors, mainly the ungulate population and browsing preference, and the browse abundance and diversity (Bohdan et al., 2020; Candaele et al., 2023; Churski et al., 2017).
However, when comparing ungulate browsing pressure between the tree species, a higher impact was observed on species such as Acer pseudoplatanus and Sorbus aucuparia in the TNP and Carpinus betulus in the RNP. According to Kupferschmid et al. (2019), densities of all species fall when annual browsing surpasses a threshold of 5%–10%. Our findings are also in line with other research (D'Aprile et al., 2020; Konôpka et al., 2018; Sample et al., 2023) which indicates that browsing pressure is especially high on palatable species.
In contrast, Fagus sylvatica and Picea abies face much lower browsing pressure within each study site than the highly palatable focal tree species. However, the actual loss of twig dry mass due to ungulate browsing is significant for these species at both study sites. This suggests that the moderate and less palatable species contribute considerably to high browse in forest ecosystems, even more than the more heavily browsed palatable species. Thus, this implies that browsing pressure is distributed across all available food resources among tree species, with ungulates exploring a broad range of feeding options regardless of their selectivity.
4.4
Post-browsing remnants: consequences for palatable tree regeneration
Despite the substantial browsing pressure on the tree saplings, the results indicate that some twig dry mass survives the browsing loss in all species, albeit to different extents. Among tree species, Picea abies retains the greatest browse after ungulate browsing in the TNP. Similarly, Fagus sylvatica retains the greatest browse in the RNP. This is likely due to the overall abundance of these species in their respective habitats (Fig. 5). On the other hand, Carpinus betulus and Acer pseudoplatanus were the least abundant, resulting in the smallest residual browse among the species. Although there are differences among forest communities, the high palatability of certain tree species results in greater browsing pressure, and significantly lower the residual browse. This suggests that such browsing pressure may hinder ecological succession, and suppress the growth of young generations following disturbance in natural ecosystems (Didion et al., 2009; Ramirez et al., 2019). Ungulates control the forest composition of regenerating trees by selectively browsing palatable broadleaved trees, leading to the competitive release of conifers (Rooney and Waller, 2003). For example, in TNP, the remaining browse from most broad-leaved deciduous species is less abundant than that of coniferous species after intense browsing. The ungulate browsing pressure may limit changes in species composition following natural disturbance and slow the transition from a historically conifer-dominated forest to one with diverse stands with deciduous species dominance.
However, the findings also show that intense browsing pressure has not eliminated the highly palatable tree species such as Carpinus betulus and Acer pseudoplatanus. Despite the significantly lower amount of browse remnants of these palatable species, their regeneration can still be found, although in most cases restricted to the lower height classes. This is because the higher abundance of potential fodder provided by less palatable focal tree species mitigates the impact of browsing pressure on palatable trees. Escaping a year of ungulate browsing could enhance survival and recruitment prospects, even though sapling recruitment success is strongly influenced by canopy gap conditions. This is particularly notable because most browsing loss on the saplings for all species, as observed in our study sites, occurs at heights below 1 m and is stronger in RNP.
5.
Conclusion
This study presents a novel approach to assess tree regeneration in forest ecosystems by measuring browse abundance and browsing pressure. A relationship was established between the browsing intensity index (BI) and ungulate browsing pressure on tree saplings, allowing the BI to estimate actual browsing pressure on different tree species. Across all species, saplings in the lower height classes exhibit the greatest browse abundance and twig dry mass loss due to ungulate browsing. The impact of ungulate browsing varies greatly between tree species within forest ecosystems. In the TNP, Acer pseudoplatanus and Sorbus aucuparia experienced higher browsing pressure than other species, while Carpinus betulus experienced more intense browsing in the RNP, resulting in a significantly lower dry mass of twigs remaining. In contrast, species such as Picea abies and Fagus sylvatica tend to retain more twigs after browsing, which increases their potential for recruitment into the forest canopy. Despite high browsing pressure, palatable species can thrive and regenerate in forest ecosystems. This resilience is due to the abundant alternative food sources provided by less palatable focal species, which mitigate the effects of ungulate browsing. Because, increasing the overall browse abundance is associated with reducing browsing pressure, particularly for highly palatable species, where this effect is more pronounced compared to less palatable species. The findings have important implications for forest management, particularly in areas where ungulates threaten tree regeneration. Increasing browse abundance may reduce browsing pressure on palatable species, promoting balanced regeneration and biodiversity. From a methodology perspective, previous studies, such as Rhodes and St. Clair (2018) and Sample et al. (2023), have linked ungulate abundance to browsing intensity, with higher browsing intensities associated with higher deer densities. In contrast, this study evaluates ungulate browsing pressure based on twig dry mass loss per species, independent of ungulate abundance. This approach provides a simpler method for estimating browsing pressure and assessing tree regeneration potential, with applications to large-scale forest ecosystem studies. Finally, we suggest a long-term study to examine the combined effects of canopy gaps and species vulnerability levels, strengthening our argument that browse abundance plays a critical role in protecting palatable species from ungulate browsing pressure in natural forest ecosystems.
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Jerzy Szwagrzyk reports financial support was provided by Polish National Science Foundation (NCN), grant No. 2018/31/B/NZ8/02786 (project title: “How do large-scale disturbances influence the relationship between ungulate herbivory and natural regeneration in temperate forests?”) from the Ministry of Science and Higher Education of the Republic of Poland within the framework of statutory activities SUB/040011-D019/2020 of the Department of Forest Biodiversity, University of Agriculture in Krakow. If there are other authors, they declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Akashi, N., Unno, A., Uno, H., 2022. The browsing ratio as an index of the impact of deer browsing on tree seedlings. Trees People 8, 100276. .
Ammer, C., 1996. Impact of ungulates on structure and dynamics of natural regeneration of mixed mountain forests in the Bavarian Alps. For. Ecol. Manag. 88 (1–2), 43–53. .
Angst, J.K., Kupferschmid, A.D., 2023. Assessing browsing impact in beech forests: the importance of tree responses after browsing. Diversity 15 (2), 262. .
Beguin, J., Tremblay, J.P., Thiffault, N., Pothier, D., Côté, S.D., 2016. Management of forest regeneration in boreal and temperate deer-forest systems: challenges, guidelines, and research gaps. Ecosphere 7 (10), e01488. .
Bernard, M., Barrere, J., Morin, X., Saïd, S., Boulanger, V., Granda, E., Benavides, R., Jactel, H., Heurich, M., Rabasa, S.G., Valladares, F., Kunstler, G., 2024. Elevation affects both the occurrence of ungulate browsing and its effect on tree seedling growth for four major tree species in European mountain forests. Ann. For. Sci. 81 (1), 13. .
Bödeker, K., Ammer, C., Knoke, T., Heurich, M., 2021a. Determining statistically robust changes in ungulate browsing pressure as a basis for adaptive wildlife management. Forests 12 (8), 1030. .
Bödeker, K., Ammer, C., Knoke, T., Heurich, M., 2021b. Determining statistically robust changes in ungulate browsing pressure as a basis for adaptive wildlife management. Forests 12 (8), 1030. .
Bödeker, K., Jordan-Fragstein, C., Vor, T., Ammer, C., Knoke, T., 2023. Abrupt height growth setbacks show overbrowsing of tree saplings, which can be reduced by raising deer harvest. Sci. Rep. 13 (1), 12021. .
Bodziarczyk, J., Szwagrzyk, J., Zwijacz-Kozica, T., Zięba, A., Szewczyk, J., Gazda, A., 2019. The structure of forest stands in the Tatra National Park: The results of 2016–2017 inventory. For. Res. Pap. 80 (1), 13–21. .
Bodziarczyk, J., Zwijacz-Kozica, T., Gazda, A., Szewczyk, J., Frączek, M., Zięba, A., Szwagrzyk, J., 2017. Species composition, elevation, and former management type affect browsing pressure on forest regeneration in the Tatra National Park. For. Res. Pap. 78 (3), 238–247. .
Bohdan, K., Pajtík, J., Bošeľa, M., Šebeň, V., Shipley, L.A., 2020. Modeling forage potential for red deer (Cervus elaphus): a tree-level approach. Eur. J. For. Res. 139 (3), 419–430. .
Borkowski, J., 2004. Distribution and habitat use by red and roe deer following a large forest fire in South-western Poland. For. Ecol. Manag. 201 (2–3), 287–293. .
Borowski, Z., Gil, W., Bartoń, K., Zajączkowski, G., Łukaszewicz, J., Tittenbrun, A., Radliński, B., 2021. Density-related effect of red deer browsing on palatable and unpalatable tree species and forest regeneration dynamics. For. Ecol. Manag. 496, 119442. .
Boulanger, V., Baltzinger, C., Saïd, S., Ballon, P., Picard, J.F., Dupouey, J.L., 2009. Ranking temperate woody species along a gradient of browsing by deer. For. Ecol. Manag. 258 (7), 1397–1406. .
Brandner, T.A., Peterson, R.O., Risenhoover, K.L., 1990. Balsam fir on isle royale: effects of moose herbivory and population density. Ecology 71 (1), 155–164. .
Candaele, R., Ligot, G., Licoppe, A., Lievens, J., Fichefet, V., Jonard, M., André, F., Lejeune, P., 2023. Interspecific growth reductions caused by wild ungulates on tree seedlings and their implications for temperate Quercus-Fagus forests. Forests 14 (7), 1330. .
Čermák, P., Horsák, P., Špiřík, M., Mrkva, R., 2009. Relationships between browsing damage and woody species dominance. J. For. Sci. 55 (1), 23–31. .
Champagne, E., Raymond, P., Royo, A.A., Speed, J.D.M., Tremblay, J.P., Côté, S.D., 2021. A review of ungulate impacts on the success of climate-adapted forest management strategies. Curr. Rep. 7 (4), 305–320. .
Churski, M., Bubnicki, J.W., Jędrzejewska, B., Kuijper, D.P.J., Cromsigt, J.P.G.M., 2017. Brown world forests: increased ungulate browsing keeps temperate trees in recruitment bottlenecks in resource hotspots. New Phytol. 214 (1), 158–168. .
Clasen, A., Somers, B., Pipkins, K., Tits, L., Segl, K., Brell, M., Kleinschmit, B., Spengler, D., Lausch, A., Förster, M., 2015. Spectral unmixing of forest crown components at close range, airborne and simulated Sentinel-2 and EnMAP spectral imaging scale. Rem. Sens. 7 (11), 15361–15387. .
D'Aprile, D., Vacchiano, G., Meloni, F., Garbarino, M., Motta, R., Ducoli, V., Partel, P., 2020. Effects of twenty years of ungulate browsing on forest regeneration at Paneveggio reserve, Italy. Forests 11 (6), 612. .
De Pauw, K., Sanczuk, P., Meeussen, C., Depauw, L., De Lombaerde, E., Govaert, S., Vanneste, T., Brunet, J., Cousins, S.A.O., Gasperini, C., Hedwall, P.O., Iacopetti, G., Lenoir, J., Plue, J., Selvi, F., Spicher, F., Uria-Diez, J., Verheyen, K., Vangansbeke, P., De Frenne, P., 2022. Forest understorey communities respond strongly to light in interaction with forest structure, but not to microclimate warming. New Phytol. 233 (1), 219–235. .
Didion, M., Kupferschmid, A.D., Bugmann, H., 2009. Long-term effects of ungulate browsing on forest composition and structure. For. Ecol. Manag. 258, S44–S55. .
Faison, E.K., DeStefano, S., Foster, D.R., Barker Plotkin, A., 2016. Functional response of ungulate browsers in disturbed eastern hemlock forests. For. Ecol. Manag. 362, 177–183. .
Fynn, R.W.S., Augustine, D.J., Fuhlendorf, S.D., 2019. Managing browsing and grazing ungulates. In: Gordon, Ⅰ., Prins, H. (Eds.), The Ecology of Browsing and Grazing Ⅱ, Ecological Studies, vol. 239. Springer, Cham, pp. 321–338. .
Gaudry, W., Gaillard, J.M., Saïd, S., Mårell, A., Baltzinger, C., Rocquencourt, A., Bonenfant, C., 2022. Population density and plant availability interplay to shape browsing intensity by roe deer in a deciduous forest. For. Ecol. Manag. 515, 120153. .
Hamberg, L., Lehvävirta, S., Kotze, D.J., Heikkinen, J., 2015. Tree species composition affects the abundance of rowan (Sorbus aucuparia L.) in urban forests in Finland. J. Environ. Manag. 151, 369–377. .
Hardalau, D., Codrean, C., Iordache, D., Fedorca, M., Ionescu, O., 2024. The expanding thread of ungulate browsing—a review of forest ecosystem effects and management approaches in Europe. Forests 15 (8), 1311. .
Heuze, P., Schnitzler, A., Klein, F., 2005. Is browsing the major factor of silver fir decline in the Vosges Mountains of France? For. Ecol. Manag. 217 (2–3), 219–228. .
Hidding, B., Tremblay, J.P., Côté, S.D., 2012. Survival and growth of balsam fir seedlings and saplings under multiple controlled ungulate densities. For. Ecol. Manag. 276, 96–103. .
Holm, J.A., Thompson, J.R., McShea, W.J., Bourg, N.A., 2013. Interactive effects of chronic deer browsing and canopy gap disturbance on forest successional dynamics. Ecosphere 4 (11), 1–23. .
Horsley, S.B., Stout, S.L., DeCalesta, D.S., 2003. White-tailed deer impact on the vegetation dynamics of a northern hardwood forest. Ecol. Appl. 13 (1), 98–118. .
Knapp, S.P., Webster, C.R., Kern, C.C., 2019. The composition and height of saplings capturing silvicultural gaps at two long-term experiments in managed northern hardwood forests. Forests 10 (10), 855. .
Konôpka, B., Pajtík, J., Shipley, L.A., 2018. Intensity of red deer browsing on young rowans differs between freshly-felled and standing individuals. For. Ecol. Manag. 429, 511–519. .
Kuijper, D.P.J., Cromsigt, J.P.G.M., Churski, M., Adam, B., Jedrzejewska, B., Jedrzejewski, W., 2009. Do ungulates preferentially feed in forest gaps in European temperate forest? For. Ecol. Manag. 258 (7), 1528–1535. .
Kupferschmid, A.D., Bütikofer, L., Hothorn, T., Schwyzer, A., Brang, P., 2019. Quantifying the relative influence of terminal shoot browsing by ungulates on tree regeneration. For. Ecol. Manag. 446, 331–344. .
Kupferschmid, A.D., Wasem, U., Bugmann, H., 2015. Browsing regime and growth response of Abies alba saplings planted along light gradients. Eur. J. For. Res. 134 (1), 75–87. .
Łukaszewicz, J., Zajączkowski, G., Gil, W., Tittenburn, A., Radliński, B., 2020. Zmiany składu gatunkowego i struktury drzewostanów w Roztoczańskim Parku Narodowym w okresie 2007−2017 [Changes in the species composition and structure of forest stands in the Roztocze National Park in the years 2007−2017]. Sylwan 164 (8), 683–694. .
Madalcho, A.B., Gazda, A., Wanic, T., Szwagrzyk, J., 2024. Influence of soil quality on the browsing intensity of ungulate herbivores on tree species in European forests. Forests 15 (4), 708. .
Madalcho, A.B., Szwagrzyk, J., Maciejewski, Z., Gazda, A., Bodziarczyk, J., 2023. Allometric scaling models for tree saplings: assessing potential fodder for ungulate herbivores. Trees Forest People 13, 100424. .
Mårell, A., Hamard, J.P., Pérot, T., Perret, S., Korboulewsky, N., 2018. The effect of deer browsing and understory light availability on stump mortality and sprout growth capacity in sessile oak. For. Ecol. Manag. 430, 134–142. .
Mclaren, B.E., Peterson, R.O., 1994. Wolves , moose , and tree rings on Isle Royale. Science 266 (5190), 1555–1558. JSTOR. . (Accessed 15 September 2024).
Nopp-Mayr, U., Reimoser, S., Reimoser, F., Sachser, F., Obermair, L., Gratzer, G., 2020. Analyzing long-term impacts of ungulate herbivory on forest-recruitment dynamics at community and species level contrasting tree densities versus maximum heights. Sci. Rep. 10 (1), 20274. .
Ohse, B., Seele, C., Holzwarth, F., Wirth, C., 2017. Different facets of tree sapling diversity influence browsing intensity by deer dependent on spatial scale. Ecol. Evol. 7 (17), 6779–6789. .
Pajtík, J., Konôpka, B., Bošel’a, M., Šebeň, V., Kaštier, P., 2015. Modelling forage potential for red deer: a case study in post-disturbance young stands of rowan. Ann. For. Res. 58 (1), 91–107. .
Petritan, A.M., von Lüpke, B., Petritan, I.C., 2009. Influence of light availability on growth, leaf morphology and plant architecture of beech (Fagus sylvatica L.), maple (Acer pseudoplatanus L.) and ash (Fraxinus excelsior L.) saplings. Eur. J. For. Res. 128 (1), 61–74. .
Pielech, R., Różański, W., Zięba, A., Zwijacz-Kozica, T., Kauzal, P., Foremnik, K., Bodziarczyk, J., Szwagrzyk, J., 2021. Forest communities of the Tatra Mountains: a classification based on a permanent plot inventory in the Tatra National Park (Poland). Tuexenia 41, 11–36. .
Pranzini, N., Maiorano, L., Cosentino, F., Thuiller, W., Santini, L., 2024. The role of species interactions in shaping the geographic pattern of ungulate abundance across African savannah. Sci. Rep. 14 (1), 19674. .
Ramirez, J.I., Jansen, P.A., den Ouden, J., Goudzwaard, L., Poorter, L., 2019. Long-term effects of wild ungulates on the structure, composition and succession of temperate forests. For. Ecol. Manag. 432, 478–488. .
Rhodes, A.C., St Clair, S.B., 2018. Measures of browse damage and indexes of ungulate abundance to quantify their impacts on aspen forest regeneration. Ecol. Indicat. 89, 648–655. .
Rooney, T.P., Waller, D.M., 2003. Direct and indirect effects of white-tailed deer in forest ecosystems. For. Ecol. Manag. 181 (1–2), 165–176. .
Samojlik, T., Kuijper, D., 2013. Grazed wood-pasture versus browsed high forests. In: Rotherham, I.D. (Ed.), Trees, Forested Landscapes and Grazing Animals: A European Perspective on Woodlands and Grazed Treescapes, first ed. Routledge, London. .
Sample, R.D., Delisle, Z.J., Pierce, J.M., Swihart, R.K., Caudell, J.N., Webster, C.R., Jenkins, M.A., 2023. Predicting and indexing ungulate browse intensity from local to regional scales. Ecol. Indicat. 154, 110564. .
Shipley, L.A., Spalinger, D.E., 1995. Influence of size and density of browse patches on intake rates and foraging decisions of young moose and white-tailed deer. Oecologia 104 (1), 112–121. .
Sproull, G.J., Quigley, M.F., Sher, A., González, E., 2015. Long-term changes in composition, diversity and distribution patterns in four herbaceous plant communities along an elevational gradient. J. Veg. Sci. 26 (3), 552–563. .
Szwagrzyk, J., Gazda, A., Muter, E., Pielech, R., Szewczyk, J., Zięba, A., Zwijacz-Kozica, T., Wiertelorz, A., Pachowicz, T., Bodziarczyk, J., 2020. Effects of species and environmental factors on browsing frequency of young trees in mountain forests affected by natural disturbances. For. Ecol. Manag. 474. .
Tremblay, J.P., Huot, J., Potvin, F., 2007. Density-related effects of deer browsing on the regeneration dynamics of boreal forests. J. Appl. Ecol. 44 (3), 552–562. .
Zywiec, M., Holeksa, J., Wesolowska, M., Szewczyk, J., Zwijacz-Kozica, T., Kapusta, P., 2013. Sorbus aucuparia regeneration in a coarse-grained spruce forest - a landscape scale. J. Veg. Sci. 24 (4), 735–743. .
Żywiec, M., Ledwoń, M., 2008. Spatial and temporal patterns of rowan (Sorbus aucuparia L.) regeneration in West Carpathian subalpine spruce forest. Plant Ecol. 194 (2), 283–291. .
Table
1.
A zero-intercept linear model for the association between browsing intensity index (BI) and the percentage of twig dry mass of a sapling eliminated by ungulate browsing. BI as a predictor variable was used to estimate the ungulate browsing pressure at each of the study sites.
Table
2.
The effect of browse abundance and species palatability level on browsing pressure of tree species. The coefficients for “low-palatability” and “moderate-palatability” show how these levels differ from the high palatability level.
Table
3.
Wilcoxon rank sum test with continuity correction was used to compare twig dry mass loss due to ungulate browsing and the browsing pressure between RNP and TNP. A Non-parametric test (Mann Whitney U) (p < 0.05).
Table
4.
Wilcoxon rank sum test with continuity correction for comparison between twig dry mass remaining after ungulate browsing in RNP and TNP. A nonparametric test (Mann Whitney U) with 95% confidence.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds