Armas, C., Pugnaire, F.I., 2005. Plant interactions govern population dynamics in a semi-arid plant community. J. Ecol. 93, 978-989. .
|
Bashirzadeh, M., Soliveres, S., Farzam, M., Ejtehadi, H., 2022. Plant-plant interactions determine taxonomic, functional and phylogenetic diversity in severe ecosystems. Global Ecol. Biogeogr. 31, 649-662. .
|
Bertness, M.D., Callaway, R., 1994. Positive interactions in communities. Trends Ecol. Evol. 9, 191-193. .
|
Bjorkman, A.D., Myers-Smith, I.H., Elmendorf, S.C., Normand, S., Rueger, N., Beck, P.S.A., Blach-Overgaard, A., Blok, D., Cornelissen, J.H.C., Forbes, B.C., Georges, D., Goetz, S.J., Guay, K.C., Henry, G.H.R., HilleRisLambers, J., Hollister, R.D., Karger, D.N., Kattge, J., Manning, P., Prevéy, J.S., Rixen, C., Schaepman-Strub, G., Thomas, H.J.D., Vellend, M., Wilmking, M., Wipf, S., Carbognani, M., Hermanutz, L., Lévesque, E., Molau, U., Petraglia, A., Soudzilovskaia, N.A., Spasojevic, M.J., Tomaselli, M., Vowles, T., Alatalo, J.M., Alexander, H.D., Anadon-Rosell, A., Angers-Blondin, S., te Beest, M., Berner, L., Björk, R.G., Buchwal, A., Buras, A., Christie, K., Cooper, E.J., Dullinger, S., Elberling, B., Eskelinen, A., Frei, E.R., Grau, O., Grogan, P., Hallinger, M., Harper, K.A., Heijmans, M.M.P.D., Hudson, J., Hülber, K., Iturrate-Garcia, M., Iversen, C.M., Jaroszynska, F., Johnstone, J.F., Jorgensen, R.H., Kaarlejärvi, E., Klady, R., Kuleza, S., Kulonen, A., Lamarque, L.J., Lantz, T., Little, C.J., Speed, J.D.M., Michelsen, A., Milbau, A., Nabe-Nielsen, J., Nielsen, S.S., Ninot, J.M., Oberbauer, S.F., Olofsson, J., Onipchenko, V.G., Rumpf, S.B., Semenchuk, P., Shetti, R., Collier, L.S., Street, L.E., Suding, K.N., Tape, K.D., Trant, A., Treier, U.A., Tremblay, J.P., Tremblay, M., Venn, S., Weijers, S., Zamin, T., Boulanger-Lapointe, N., Gould, W.A., Hik, D.S., Hofgaard, A., Jónsdóttir, I.S., Jorgenson, J., Klein, J., Magnusson, B., Tweedie, C., Wookey, P.A., Bahn, M., Blonder, B., van Bodegom, P.M., Bond-Lamberty, B., Campetella, G., Cerabolini, B.E.L., Chapin, F.S., Cornwell, W.K., Craine, J., Dainese, M., de Vries, F.T., Díaz, S., Enquist, B.J., Green, W., Milla, R., Niinemets, Ü., Onoda, Y., Ordoñez, J.C., Ozinga, W.A., Penuelas, J., Poorter, H., Poschlod, P., Reich, P.B., Sande, B., Schamp, B., Sheremetev, S., Weiher, E., 2018. Plant functional trait change across a warming tundra biome. Nature 562, 57-62. .
|
Bruno, J.F., Stachowicz, J.J., Bertness, M.D., 2003. Inclusion of facilitation into ecological theory. Trends Ecol. Evol. 18, 119-125. .
|
Buchwal, A., Weijers, S., Blok, D., Elberling, B., 2019. Temperature sensitivity of willow dwarf shrub growth from two distinct High Arctic sites. Int. J. Biometeorol. 63, 167-181. .
|
Büntgen, U., Hellmann, L., Tegel, W., Normand, S., Myers-Smith, I.H., Kirdyanov, A.V., Nievergelt, D., Schweingruber, F.H., 2015. Temperature-induced recruitment pulses of Arctic dwarf shrub communities. J. Ecol. 103, 489-501. .
|
Butterfield, B.J., Betancourt, J.L., Turner, R.M., Briggs, J.M., 2010. Facilitation drives 65 years of vegetation change in the Sonoran Desert. Ecology 91, 1132-1139. .
|
Callaway, R.M., Brooker, R.W., Choler, P., Kikvidze, Z., Lortie, C.J., Lortie, R., Michalet, R., Paolini, L., Pugnaire, F.I., Newingham, B., Aschehoug, E.T., Armas, C., Kikodze, D., Cook, B.J., 2002. Positive interactions among alpine plants increases with stress. Nature 417, 844-848. .
|
Camarero, J.J., Gutiérrez, E., 2004. Pace and pattern of recent treeline dynamics: response of ecotones to climatic variability in the Spanish Pyrenees. Clim. Change 63, 181-200. .
|
Cameron, H., Coulson, T., Marshall, D.J., 2019. Size and density mediate transitions between competition and facilitation. Ecol. Lett. 22, 1879-1888. .
|
Cavieres, L.A., Hernández-Fuentes, C., Sierra-Almeida, A., Kikvidze, Z., 2016. Facilitation among plants as an insurance policy for diversity in Alpine communities. Funct. Ecol. 30, 52-59. .
|
Chen, J., He, X., Wang, S., Yang, Y., Sun, H., 2019. Cushion and shrub ecosystem engineers contribute differently to diversity and functions in alpine ecosystems. J. Veg. Sci., 362-374. .
|
Chen, J., Yang, Y., Wang, S., Sun, H., Schöb, C., 2020. Shrub facilitation promotes selective tree establishment beyond the climatic treeline. Sci. Total Environ. 708, 134618. .
|
Choler, P., Michalet, R., Callaway, R.M., 2001. Facilitation and competition on gradients in alpine plant communities. Ecology 82, 3295-3308. .
|
Conti, G., Gorné, L.D., Zeballos, S.R., Lipoma, M.L., Gatica, G., Kowaljow, E., Valiente-Banuet, J.I., Cuchietti, A., Poca, M., Pestoni, S., Fernandes, P.M., 2019. Developing allometric models to predict the individual aboveground biomass of shrubs worldwide. Global Ecol. Biogeogr. 28, 961-975. .
|
|
Cook, E.R., Kairiukstis, L.A., 1990. Methods of Dendrochronology: Applications in the Environmental Sciences. Kluwer, Dordrecht, The Netherlands.
|
Devi, N.M., Kukarskih, V.V., Galimova, A.A., Mazepa, V.S., Grigoriev, A.A., 2020. Climate change evidence in tree growth and stand productivity at the upper treeline ecotone in the Polar Ural Mountains. For. Ecosyst. 7, 7. .
|
Elliott, G.P., 2011. Influences of 20th century warming at the upper tree line contingent on local- scale interactions: evidence from a latitudinal gradient in the Rocky Mountains, USA. Global Ecol. Biogeogr. 20, 46-57. .
|
Enquist, B.J., West, G.B., Brown, J.H., 2009. Extensions and evaluations of a general quantitative theory of forest structure and dynamics. Proc. Natl. Acad. Sci. U.S.A. 106, 7046-7051. .
|
Ettinger, A., HilleRisLambers, J., 2017. Competition and facilitation may lead to asymmetric range shift dynamics with climate change. Global Change Biol. 23, 3921-3933. .
|
Fajardo, A., McIntire, E.J.B., 2011. Under strong niche overlap conspecifics do not compete but help each other to survive: facilitation at the intraspecific level. J. Ecol. 99, 642-650. .
|
Fang, J., Wang, Z., Tang, Z., 2011. Atlas of Woody Plants in China: Distribution and
Climate. Springer, Berlin, Heidelberg. doi: .
|
Fichtner, A., Härdtle, W., Li, Y., Bruelheide, H., Kunz, M., von Oheimb, G., 2017. From competition to facilitation: how tree species respond to neighbourhood diversity. Ecol. Lett. 20, 892-900. .
|
Filazzola, A., Lortie, C.J., 2014. A systematic review and conceptual framework for the mechanistic pathways of nurse plants. Global Ecol. Biogeogr. 23, 1335-1345. .
|
Gao, S., Liang, E.Y., Liu, R., Babst, F., Camarero, J.J., Fu, Y.S.H., Piao, S., Rossi, S., Shen, M., Wang, T., Peñuelas, J., 2022. An earlier start of the thermal growing season enhances tree growth in cold humid areas but not in dry areas. Nat. Ecol. Evol. 6, 397-404. .
|
García-Cervigón, A.I., Linares, J.C., Aibar, P., Olano, J.M., 2015. Facilitation promotes changes in leaf economics traits of a perennial forb. Oecologia 179, 103-116. .
|
Grau, O., Ninot, J.M., Blanco-Moreno, J.M., van Logtestij, R.S.P., Cornelissen, J.H.C., Callaghan, T.V., 2012. Shrub-tree interactions and environmental changes drive treeline dynamics in the Subarctic. Oikos 121, 1680-1690. .
|
Han, Y., Wang, Y., Liu, B., Huang, R., Camarero, J.J., 2022. Moisture mediates temperature-growth couplings of high-elevation shrubs in the Tibetan plateau. Trees Struct. Funct. 36, 273-281. .
|
Harper, K.A., Danby, R.K., De Fields, D.L., Lewis, K.P., Trant, A.J., Starzomski, B.M., Savidge, R., Hermanutz, L., 2011. Tree spatial pattern within the forest–tundra ecotone: a comparison of sites across Canada. Can. J. For. Res. 41, 479-489. .
|
He, Q., Bertness, M.D., 2014. Extreme stresses, niches, and positive species interactions along stress gradients. Ecology 95, 1437-1443. .
|
He, Q., Bertness, M.D., Altieri, A.H., 2013. Global shifts towards positive species interactions with increasing environmental stress. Ecol. Lett. 16, 695-706. .
|
Holmgren, M., Scheffer, M., 2010. Strong facilitation in mild environments: the stress gradient hypothesis revisited. J. Ecol. 98, 1269-1275. .
|
IPCC, 2022. Climate Change 2022: Impacts, Adaptation and Vulnerability. Contribution
of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel
on Climate Change. . Cambridge, UK and
New York, USA.
|
Kikvidze, Z., Pugnaire, F.I., Brooker, R.W., Choler, P., Lortie, C.J., Michalet, R., Callaway, R.M., 2005. Linking patterns and processes in alpine plant communities: a global study. Ecology 86, 1395-1400. .
|
Körner, C., 2012a. Alpine Treelines: Functional Ecology of the Global High Elevation Tree
Limits. Springer, Basel. .
|
Körner, C., 2012. Treelines will be understood once the functional difference between a tree and a shrub is. Ambio 41, 197-206. .
|
Lara-Romero, C., de la Cruz, M., Escribano-Ávila, G., García-Fernández, A., Iriondo, J.M., 2016. What causes conspecific plant aggregation? Disentangling the role of dispersal, habitat heterogeneity and plant-plant interactions. Oikos 125, 1304-1313. https://doi.org/10.1111/jvs.12363.
|
Law, R., Illian, J., Burslem, D.F.R.P., Gratzer, G., Gunatilleke, C.V.S., Gunatilleke, I.A.U.N., 2009. Ecological information from spatial patterns of plants: insights from point process theory. J. Ecol. 97, 618-628. .
|
Lefcheck, J.S., 2016. PIECEWISESEM: piecewise structural equation modelling in R for ecology, evolution and systematics. Methods Ecol. Evol. 7, 573-579. .
|
Li, J.,, Okin, G.S., Blackwell, J., 2017. Abiotic processes are insufficient for fertile island development: a 10-year artificial shrub experiment in a desert grassland. Geophys. Res. Lett. 44, 2245-2253. .
|
Li, Q., Michalet, R., Guo, X., Xie, H., He, M., 2021. Variation in biomass and nutrients allocation of Corydalis hendersonii on the Tibetan Plateau with increasing rainfall continentality and altitude. Ecol. Indicat. 132, 108244. .
|
Li, Q., Shen, X., Huang, Q., Sun, F., Zhou, J., Ma, X., Ran, Z., Chen, Y., Li, Z., Yan, Y., Zhang, X., Gao, W., Liu, L., 2021. Resource islands of Salix cupularis facilitating seedling emergence of the companion herbs in the restoration process of decertified alpine meadow, the Tibetan Plateau. J. Environ. Manag. 289, 112434. .
|
Li, X., Liang, E.Y., Camarero, J.J., Rossi, S., Zhang, J., Zhu, H., Fu, Y.S.H., Sun, J., Wang, T., Piao, T., Peñuelas, J., 2023. Warming-induced phenological mismatch between trees and shrubs explains high-elevation forest expansion. Natl. Sci. Rev. 10, nwad182. .
|
Liang, E.Y., Wang, Y., Piao, S., Lu, X., Camarero, J.J., Zhu, H., Zhu, L., Ellison, A.M., Ciais, P., Peňuelas, J., 2016. Species interactions slow warming-induced upward shifts of treelines on the Tibetan Plateau. Proc. Natl. Acad. Sci. U.S.A. 113, 4380-4385. .
|
Liu, Q., Peng, C., Schneider, R., Cyr, D., Liu, Z., Zhou, X., Du, M., Li, P., Jiang, Z., McDowell, N.G., Kneeshaw, D., 2023. Vegetation browning: global drivers, impacts, and feedbacks. Trends Plant Sci. 28, 1014-1032. .
|
López, R.P., Squeo, F.A., Armas, C., Kelt, D.A., Gutiérrez, J.R., 2016. Enhanced facilitation at the extreme end of the aridity gradient in the Atacama Desert: a community- level approach. Ecology 97, 1593-1604. .
|
Lu, X., Hu, F., Liang, E.Y., Sigdel, S.R., Shang, Z., Camarero, J.J., 2023. Loss of growth resilience towards the alpine shrubline. For. Ecol. Manag. 539, 121013. .
|
Lu, X., Liang, E.Y., Babst, F., Camarero, J.J., Büntgen, U., 2022. Warming-induced tipping points of Arctic and alpine shrub recruitment. Proc. Natl. Acad. Sci. U.S.A. 119, e2118120119. .
|
Lu, X., Liang, E.Y., Camarero, J.J., Ellison, A.M., 2021. An unusually high shrubline on the Tibetan Plateau. Ecology 102, e03310. .
|
Lyu, L., Zhang, Q., Deng, X., Mäkinen, H., 2016. Fine-scale distribution of treeline trees and the nurse plant facilitation on the eastern Tibetan Plateau. Ecol. Indicat. 66, 251-258. .
|
Májeková, M., de Bello, F., Dolezal, J., Lepš, J., 2014. Plant functional traits as determinants of population stability. Ecology 95, 2369-2374. .
|
Michalet, R., Le Bagousse-Pinguet, Y., Maalouf, J.P., Lortie, C.J., 2014. Two alternatives to the stress-gradient hypothesis at the edge of life: the collapse of facilitation and the switch from facilitation to competition. J. Veg. Sci. 25, 609-613. .
|
Michalet, R., Brooker, R.W., Cavieres, L.A., Kikvidze, Z., Lortie, C.J., Pugnaire, F.I., Valiente-Banuet, A., Callaway, R.M., 2006. Do biotic interactions shape both sides of the humped-back model of species richness in plant communities? Ecol. Lett. 9, 767-773. .
|
Michalet, R., Choler, P., Callaway, R.M., Whitham, T.G., 2021. Rainfall continentality, via the winter Gams angle, provides a new dimension to biogeographical distributions in the western United States. Global Ecol. Biogeogr. 30, 384-397. .
|
Mihoč, M.A.K., Giménez-Benavides, L., Pescador, D.S., Sánchez, A.M., Cavieres, L.A., Escudero, A., 2016. Soil under nurse plants is always better than outside: a survey on soil amelioration by a complete guild of nurse plants across a long environmental gradient. Plant Soil 408, 31-41. .
|
Muhamed, H., Touzard, B., Le Bagousse-Pinguet, Y., Michalet, R., 2013. The role of biotic interactions for the early establishment of oak seedlings in coastal dune forest communities. For. Ecol. Manag. 297, 67-74. .
|
Myers-Smith, I.H., Hik, D.S., 2018. Climate warming as a driver of tundra shrubline advance. J. Ecol. 106, 547-560. .
|
Nathan, R., Katul, G.G., Horn, H.S., Thomas, S.M., Oren, R., Avissar, R., Pacala, S.W., Levin, S.A., 2002. Mechanisms of long-distance dispersal of seeds by wind. Nature 418, 409-413. .
|
Pausas, J.G., Bond, W.J., 2022. Feedbacks in ecology and evolution. Trends Ecol. Evol. 37, 637-644. .
|
Piao, S., Wang, X., Park, T., Chen, C., Lian, X., He, Y., Bjerke, J.W., Chen, A., Ciais, P., Tømmervik, H., Nemani, R.R., Myngeni, R.B., 2020. Characteristics, drivers and feedbacks of global greening. Nat. Rev. Earth Environ. 1, 14-27. .
|
Poorter, L., Bongers, L., Bongers, F., 2006. Architecture of 54 moist-forest tree species: traits, trade-offs, and functional groups. Ecology 87, 1289-1301. .
|
Rietkerk, M., Van de Koppel, J., 2008. Regular pattern formation in real ecosystems. Trends Ecol. Evol. 23, 169-175. .
|
Shen, W., Zhang, L., Gao, Y., Luo, T., 2018. Causes for treeline stability under climate warming: evidence from seed and seedling transplant experiments in southeast Tibet. For. Ecol. Manag. 408, 45-53. .
|
Shi, H., Zhou, Q., He, R., Zhang, Q., Dang, H., 2022. Climate warming will widen the lagging gap of global treeline shift relative to densification. Agr. For. Meteorol. 318, 108917. .
|
Sigdel, S.R., Liang, E.Y., Wang, Y.F., Dawadi, B., Camarero, J.J., 2020. Tree-to-tree interactions slow down Himalayan treeline shifts as inferred from tree spatial patterns. J. Biogeogr. 47, 1816-1826. .
|
Sigdel, S.R., Pandey, J., Liang, E.Y., Muhammad, S., Babst, F., Leavitt, S.W., Shen, M., Zhu, H., Salerno, F., Piao, S., Camarero, J.J., Peñuelas, J., 2021. No benefits from warming even for subnival vegetation in the central Himalayas. Sci. Bull. 66, 1825-1829. .
|
Silliman, B.R., Schrack, E., He, Q., Cope, R., Santoni, A., van der Heide, T., Jacobi, R., Jacobi, M., van de Koppel, J., 2015. Facilitation shifts paradigms and can amplify coastal restoration efforts. Proc. Natl. Acad. Sci. U.S.A. 112, 14295-14300. .
|
Trautz, A.C., Illangasekare, T.H., Rodriguez-Iturbe, I., 2017. Role of co-occurring competition and facilitation in plant spacing hydrodynamics in water-limited environments. Proc. Natl. Acad. Sci. U.S.A. 114, 9379-9384. .
|
Vega-Álvarez, J., García-Rodríguez, J.A., Cayuela, L., 2019. Facilitation beyond species richness. J. Ecol. 107, 722-734. .
|
Verdú, M., García-Fayos, P., 1996. Nucleation processes in a mediterranean bird-dispersed plant. Funct. Ecol. 10, 275-280. .
|
Verdú, M., Gómez, J.M., Valiente-Banuet, A., Schöb, C., 2021. Facilitation and plant phenotypic evolution. Trends Plant Sci. 26, 913-923. .
|
Wang, X.T., Michalet, R., Meng, L., Zhou, X., Chen, S., Du, G., Xiao, S., 2021. Direct and indirect facilitation affect community productivity through changes in functional diversity in an alpine system. Ann. Bot. 127, 241-249. .
|
Wang, Y., Liang, E.Y., Ellison, A.M., Lu, X., Camarero, J.J., 2015. Facilitation stabilizes moisture-controlled alpine juniper shrublines in the central Tibetan Plateau. Global Planet. Change 132, 20-30. .
|
Wang, Y., Liang, E.Y., Lu, X., Camarero, J.J., Babst, F., Shen, M., Peñuelas, J., 2021. Warming-induced shrubline advance stalled by moisture limitation on the Tibetan Plateau. Ecography 44, 1631-1641. .
|
Wang, Y., Pederson, N., Ellison, A.M., Buckley, H.L., Case, B.S., Liang, E.Y., Camarero, J.J., 2016. Increased stem density and competition may diminish the positive effects of warming at alpine treeline. Ecology 97, 1668-1679. .
|
Wieczorek, M., Kruse, S., Epp, L.S., Kolmogorov, A., Nikolaev, A.N., Heinrich, I., Jeltsch, F., Pestryakova, L.A., Zibulski, R., Herzschuh, U., 2017. Dissimilar responses of larch stands in northern Siberia to increasing temperatures—a field and simulation based study. Ecology 98, 2343-2355. .
|
Wiegand, T., Moloney, K.A., 2004. Rings, circles and null-models for point pattern analysis in ecology. Oikos 104, 209-229. .
|
Wiegand, T., Moloney, K.A., 2014. Handbook of Spatial Point-Pattern Analysis in Ecology.
Chapman & Hall/CRC, Boca Raton, Florida, USA. .
|
Wright, A., Schnitzer, S.A., Reich, P.B., 2014. Living close to your neighbors: the importance of both competition and facilitation in plant communities. Ecology 95, 2213-2223. .
|
Wright, A.J., Barry, K.E., Lortie, C.J., Callaway, R.M., 2021. Biodiversity and ecosystem functioning: have our experiments and indices been underestimating the role of facilitation? J. Ecol. 109, 1962-1968. .
|
Yao, T., Thompson, L., Yang, W., Yu, W.S., Gao, Y., Guo, X., Yang, X., Duan, K., Zhao, H., Xu, B., Pu, J., Lu, A., Xiang, Y., Kattel, D.B., Joswiak, D., 2012. Different glacier status with atmospheric circulations in Tibetan Plateau and surroundings. Nat. Clim. Change 2, 663-667. https://doi.org/10.1111/1365-2745.13665.
|

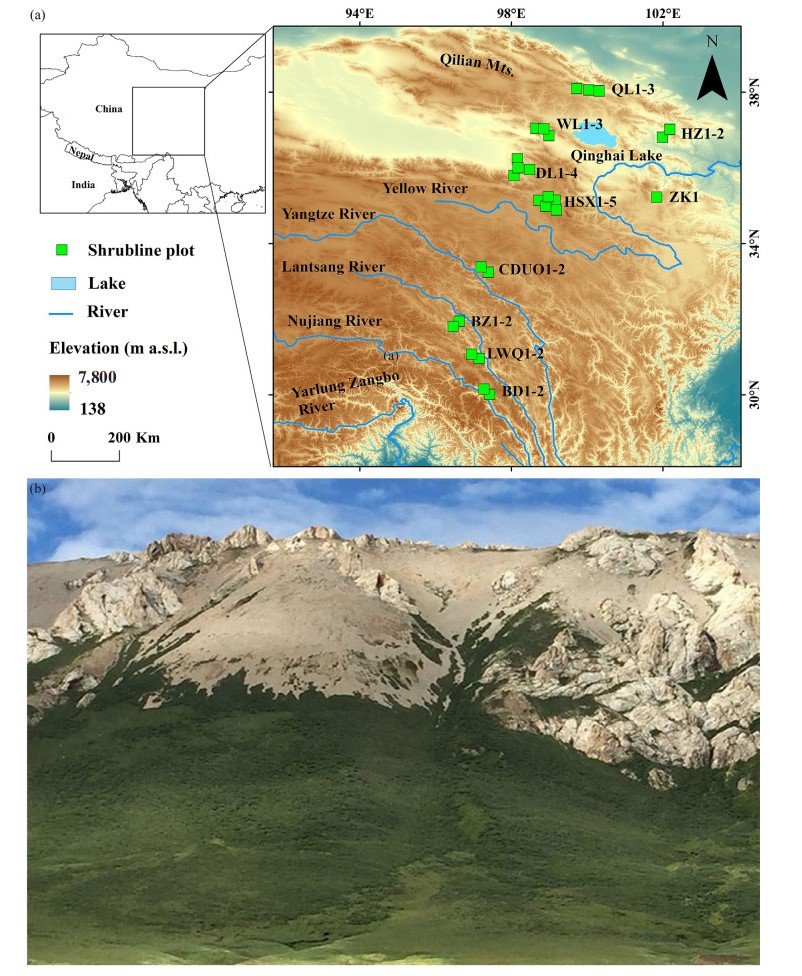
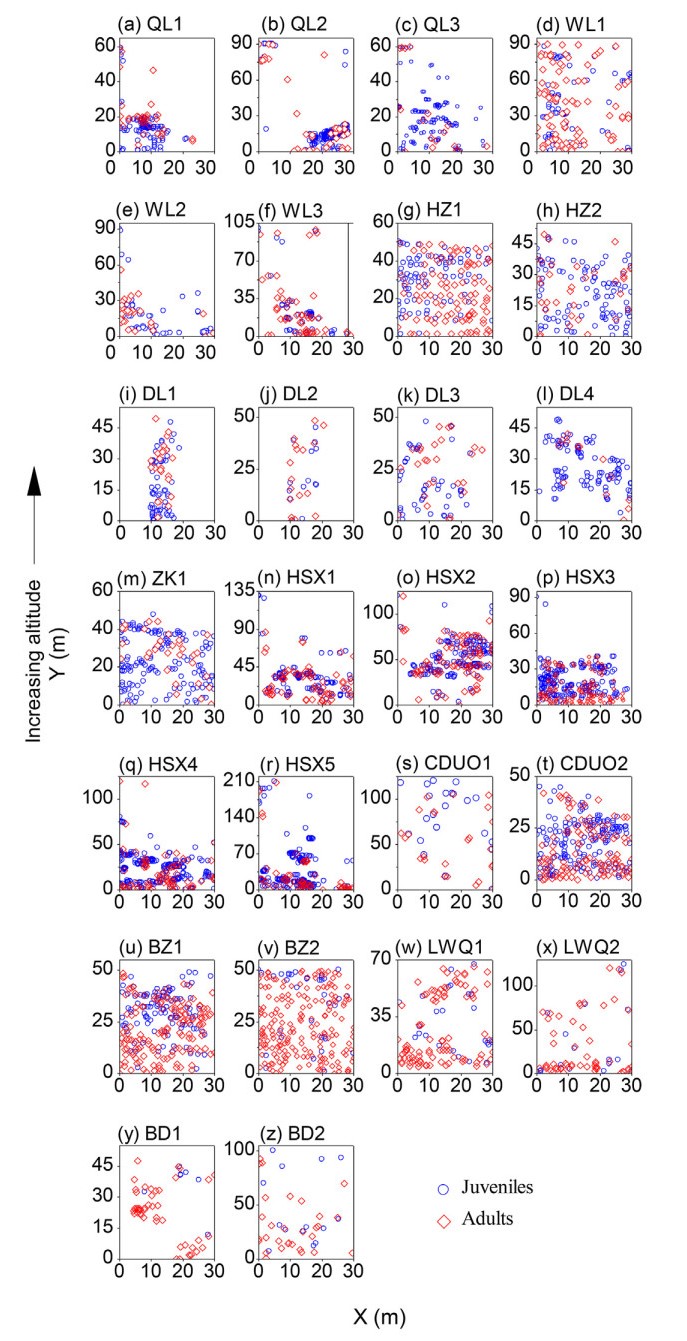
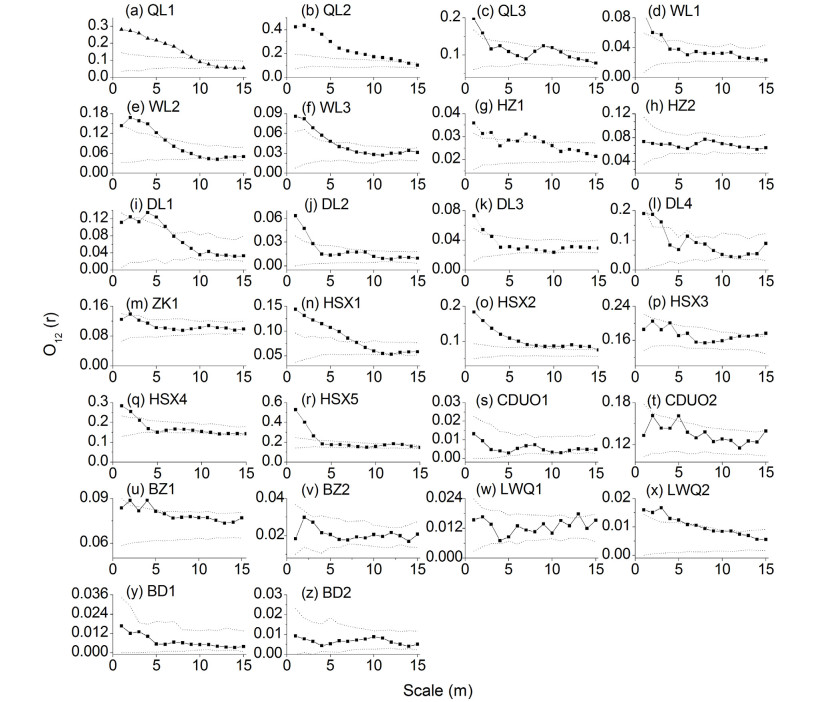
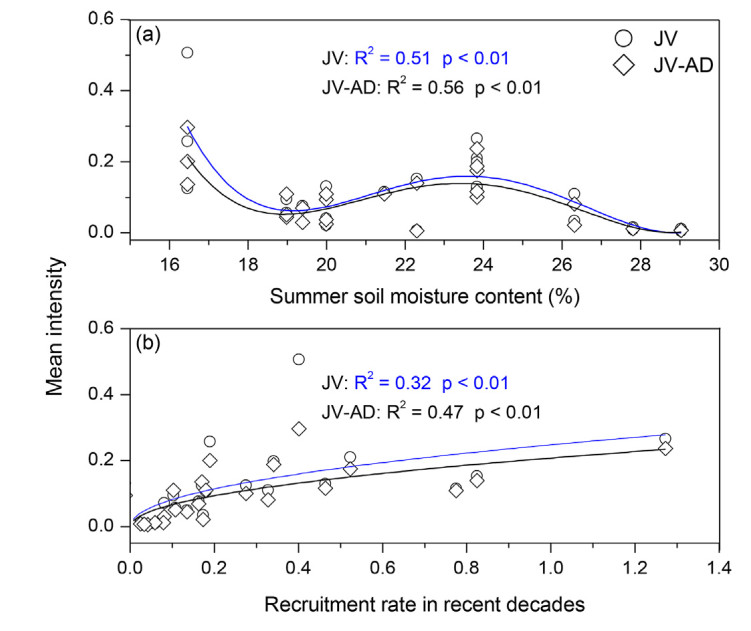
 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds