College of Forestry, Northwest A&F University, Yangling, Shaanxi, 712100, China
Funds: This work was supported by Natural Science Basic Research Program of Shaanxi (2021JQ-155), National Natural Science Foundation of China (32101511) and Chinese Universities Scientific Fund (2452020137 and 2452021073)
Background Chinese pine (Pinus tabuliformis Carr.) is one of the major afforestation species in northern China and plays a key role in restoring forest ecosystems and preserving soil and water. However, most Chinese pine plantations are experiencing ecological problems such as the low diversity of understory plants and difficulty in natural regeneration. Thinning has been widely used to maintain and improve a variety of forest ecosystem services from plantations. To date, however, few studies have been conducted to systematically determine the effects of thinning on understory plant diversity and the regeneration of Chinese pine in plantations.
Methods We conducted a literature search, and selected 22 publications covering a total of 83 treatments related to thinning effects on the species richness of understory plants and 15 publications covering a total of 43 treatments related to thinning effects on the regeneration of Chinese pine, in tree plantations of northern China. The data from the literature were synthesized and evaluated with meta-analysis approach to determine the treatment effects.
Results Compared with the control stands, thinning increased the species richness of shrubs and herbs by an average of 25.3% and 26.5%, respectively. While the varying thinning intensities all had significantly positive effects on the species richness of understory plants, only moderate thinning (30%–50%) had a positive effect on the density of regenerating seedlings and saplings of Chinese pine (60.2%). The species richness of understory plants was greatest after 14 years of thinning with an increase of 36.3%, whereas the density of regenerating Chinese pine seedlings and saplings reached a maximum after ≥11 years of thinning with an increase of 76.5%, compared to that of the unthinned stands. Thinning in the half-mature plantations had the greatest effects on the understory shrub richness (44.1%) and the density of regenerating Chinese pine seedlings and saplings (86.5%). Both single and multiple thinning were found to significantly promote the species richness of understory plants and the density of regenerating Chinese pine seedlings and saplings, and the positive effects of thinning were greater in areas with a humidity index (HI) < 30 than in areas with an HI ≥ 30. In general, age group, planting density and recovery time were prominent factors affecting the species richness of understory plants, whereas the slope, HI and recovery time were the dominant controls of the density of regenerating Chinese pine seedlings and saplings, indicating differential effects of thinning on the species richness of understory plants and the regeneration capacity of Chinese pine in plantations.
Conclusion Thinning appears to be a feasible management measure to improve the understory plant diversity and regeneration capacity of Chinese pine in plantations. We postulate that moderate thinning in half-mature forest stands with an HI < 30 can help effectively promote the species diversity of understory plants and the natural regeneration of Chinese pine, thereby maintaining a more resilient stand structure and the development of Chinese pine plantations.
As an important component of terrestrial ecosystems, plantations can not only provide wood and non-wood forest products, but also make great contributions to the reconstruction and restoration of forest ecosystems and the increase in forest carbon sinks (Pan et al., 2011; Zhang et al., 2018). Since 1990, plantation areas have added 123 million hectares worldwide, currently accounting for approximately 7% of the global forest area (FAO, 2020). To meet the demands for wood products and ecological restoration, the Chinese government launched a series of large-scale ecological restoration projects starting in the 1960s, including the “Grain for Green Project” and “Three-North Shelterbelt Program”. At present, plantation areas have reached 80 million hectares, which is approximately 36% of the total forest coverage in China (State Forestry Administration, 2018). Undoubtedly, plantations have significantly increased forest coverage and improved the regional ecological environment in the short term. However, plantations are experiencing a series of major ecological problems, such as the outbreak of diseases and pests and difficulties in natural regeneration, in the long run due to the single tree species structure and high stand density (Liu et al., 2018). Understory plants are important plant communities in plantation ecosystems. While improving soil structure and maintaining soil fertility, they also act as filters in determining the future tree species composition, which can promote the succession of plantations to a more resilient stand structure (Nilsson and Wardle, 2005; Swanson et al., 2011). Natural regeneration, as an effective way to increase the diversity of understory plants, can make full use of the self-reproductive capacity of plants to gradually transform pure plantations into heterogeneous multilayer forests or mixed forests in the process of near-natural management, thereby ensuring the spatial and temporal continuity of forest cover and sustainable management (Wang et al., 2019a). Therefore, knowing what kind of silvicultural measures should be adopted to improve the species diversity of understory plants and the natural regeneration capacity of plantations needed to alleviate the abovementioned problems are urgent issues to be solved in current forestry research.
Thinning is one of the most widely used management practices globally, and can cause a variety of ecological responses in understory vegetation (Aun et al., 2021). First, thinning increases the available space for forestland. This released ecological space is not only conducive to the survival and growth of more understory plants but also enlarges the complementary effect of the niche, which creates a suitable environment for seed germination and seedling establishment (Jiménez et al., 2015; Wang et al., 2019b). Second, thinning improves the light environment in forests by reducing stand density and increasing soil water infiltration and runoff interception, thereby alleviating moisture stress and competition for limited resources between understory plants (Hale, 2003; Trentini et al., 2017; del Campo et al., 2019). In addition, the increased soil temperature after thinning can accelerate the decomposition of litter by soil microorganisms and return nutrients to the soil (Dupuy and Chazdon, 2008). In particular, the decomposition of litter directly reduces the thickness of litter and greatly increases the effective contact between seeds and soil, which creates opportunities for seed germination and the successful establishment of seedlings (Han et al., 2014).
However, some studies have shown that thinning might lead to intense competition among understory plants due to the increased dominance of one or a few plant species, resulting in a reduction in understory species diversity and natural regeneration density (Ito et al., 2006; Sabo et al., 2009; Seiwa et al., 2021). In addition, thinning intensity, recovery time after thinning and thinning frequency can cause different responses of understory plants to thinning in different climatic conditions and stands with different successional stages (Ma et al., 2007; Gao et al., 2013; Dang et al., 2018; Gong et al., 2021). For example, Wang et al. (2019c) found that the species richness of understory plants increased with increasing thinning intensity (30%–70%) in a Chinese fir (Cunninghamia lanceolata (Lamb.) Hook.) plantation and suggested that thinning with 50% intensity could be an effective strategy to maintain the structural and functional features of its plantation ecosystems. However, Raymond et al. (2018) found that moderate thinning (31%) was the optimal intensity to promote the diversity of understory plants and the natural regeneration of target tree species by implementing different thinning intensities (20%–42%) in temperate artificial mixed forests. In addition, some studies have shown that the stability of the original ecological structure may be damaged to a large extent by the interference of different thinning frequencies, and it is difficult for these ecosystems to recover in a short period (Seiwa et al., 2021). For example, Goodwin et al. (2018) found that thinning reduced litter thickness and increased woodland availability in the early stage, but these effects were short-lived in the absence of subsequent interferences. After thinning in different forest succession stages, due to the different light and heat conditions and the availability of woodland space, the diversity of understory vegetation will also vary with community structure and tree species composition (Aavik et al., 2009). Similarly, differences in climatic conditions, such as temperature and rainfall, also profoundly determine the distribution and growth of understory plants after thinning (Ali et al., 2019). Therefore, it is essential to reveal the general law of the effects of thinning on the species diversity of understory plants and natural regeneration density and to clarify their main influencing factors.
As an endemic tree species in China, Chinese pine (Pinus tabuliformis Carr.) is one of the constructive conifer species in northern China and is often used in the construction of the ecological environment because of its drought tolerance, barren resistance and excellent soil and water conservation (Xu, 1993; Zhong et al., 2019). Chinese pine can mix with some broadleaf trees (such as Populus, Betula, Quercus) to form a good stand structure and be treated as the main timber species. However, most Chinese pine plantations planted by the government in the 1960s–1970s are mature or nearly mature, and they have experienced slow growth, pest and disease outbreaks, and degradation following fertility declines in recent decades (Wang et al., 2017). To date, many studies have reported the response of understory plants and/or natural regeneration to thinning in plantations. For example, An et al. (2012) found that the species richness of understory plants first increased and then decreased with increasing thinning intensity (46.6%–67.1%), and the number of plant species in the stands with strong thinning intensity was lower than that in the control stands. However, Dang et al. (2018) found that the understory plant richness in a Chinese pine plantation was significantly improved after 11 years of thinning (15%–45%) compared to that in control stands. Similarly, Chen and Cao (2014) found that the species richness of shrubs showed an upward trend with increasing thinning intensity (31%–65%) in a Chinese pine plantation after thinning for 30 years, but no significant difference was observed among thinning intensities (including the control treatment).
At present, the specific responses of understory plant richness and the density of regenerating seedlings and saplings to thinning in Chinese pine plantations have not been quantified in northern China, and whether there is a general pattern of responses to thinning at a large regional scale remains unclear. Therefore, at the northern scale of China, the main objectives of this study were to (1) analyze and quantify the effects of thinning on the species richness of understory plants and the density of regenerating Chinese pine seedlings and saplings; (2) reveal the influence degree and regularity of different thinning intensities, recovery time after thinning, thinning frequencies, age groups and climatic conditions on thinning; and (3) clarify the key factors affecting the species richness of understory plants and the density of regenerating seedlings and saplings in Chinese pine plantations.
2.
Materials and methods
2.1
Data extraction
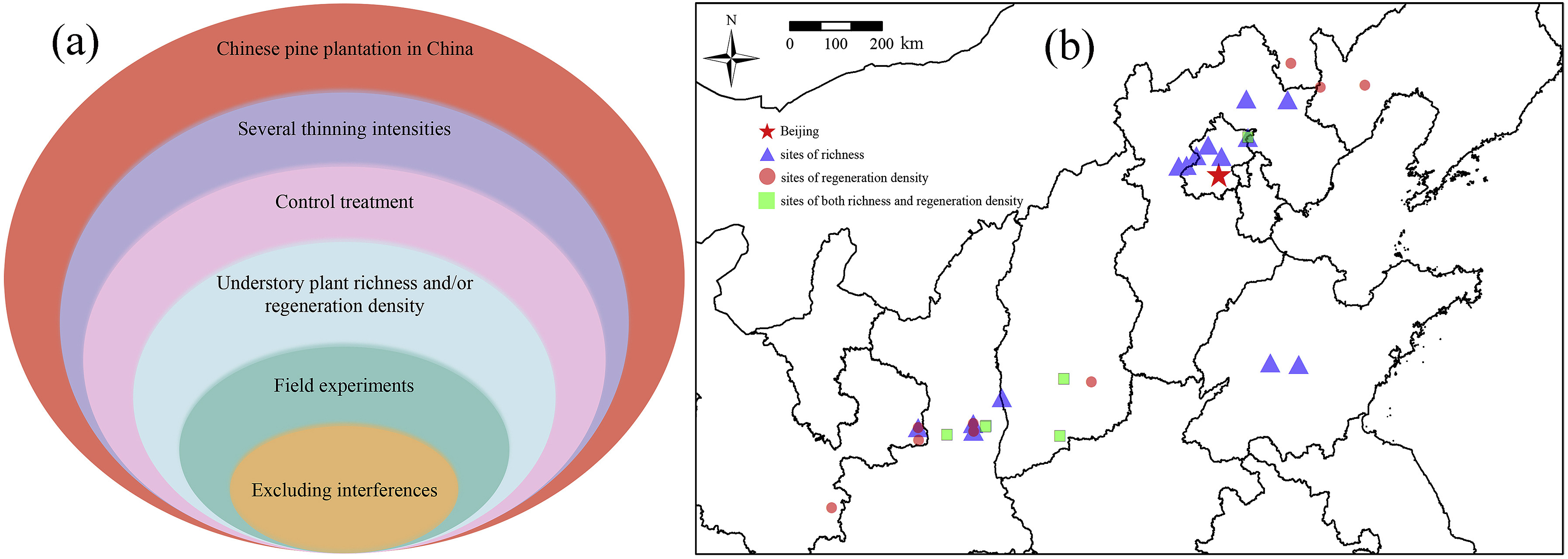
We used the online databases of Web of Science and China National Knowledge Infrastructure (CNKI) to search for peer-reviewed publications related to the “effects of thinning on the species diversity of understory plants and/or natural regeneration in Chinese pine plantations” published before October 2021. During the retrieval process, we first used different keywords or their combinations, such as “thinning, forest management, harvest, forest tending, Chinese pine plantation, natural regeneration, diversity or richness”, for the literature search. Then, we evaluated the quality of these collected studies to avoid deviations in the results. Finally, based on the criteria in Fig. 1a, 22 publications covering a total of 83 treatments related to thinning effects on the species richness of understory plants and 15 publications covering a total of 43 treatments related to thinning effects on the density of regenerating Chinese pine seedlings and saplings were screened (Table S1). As a result, our study area spanned northern China (Fig. 1b).
Figure
1.
The criteria for literature selection (a) and distribution of study sites (b). We selected articles on the effects of different intensities of thinning, which were performed without other interference in the field and included a control treatment, on the species richness of understory plants and/or the density of regenerating seedlings and saplings in Chinese pine plantations.
Information on environmental factors such as geographic coordinates, mean annual temperature (MAT), mean annual precipitation (MAP), aspect, slope, elevation, planting density, retention density, sample area, stand age or age groups at thinning, thinning frequency and recovery time were extracted from the final selected articles. The recovery time refers to the time from thinning to data investigation in the corresponding article. If multiple thinning were conducted in the same forest stand, the effect of each thinning was analyzed by regarding each as independent data for collection. When counting the thinning frequency, many articles introduced only the thinning frequency in the history of forest management without specifying the corresponding intensity of each thinning. Therefore, in the categorical variables of thinning frequency, we defined the thinning intensity as the intensity of the last thinning in the tested stands.
In this study, we mainly employed species richness (which refers to the number of species) to indicate the responses of understory plants to thinning because it is a simple and accessible indicator that can be used to uniformly measure the effects of thinning in large scale regions (Li et al., 2020), although it does not always reflect the real changes in plant communities (Wilsey et al., 2005; Mori et al., 2013). Additionally, some studies have used the number of species (S) to represent species richness, while others have used the species richness index of Gleason (G). To make the analysis easier, G was converted into S (S = G × lnA, where S refers to the number of species and A refers to the sample area). Because some screened articles classified the seedlings and saplings of understory trees as shrubs, we focused mainly on the species richness of understory shrubs (including understory shrubs and trees) and herbs. The unified unit of the density of regenerating Chinese pine seedlings and saplings was “stems·m−2”. Due to the significant positive correlation between MAT and MAP in China, the humidity index (HI = MAP/(MAT + 10)) was used to represent different climatic conditions (Deng and Shangguan, 2017).
2.2
Data structure
Considering that understory plants and natural regeneration play different roles in maintaining the stability of plantation ecosystems and that there are differences in data sets and some of the effective information that can be collected between them, in this study, the responses of the species richness of understory plants and the density of regenerating Chinese pine seedlings and saplings to thinning were divided into different data structures (Table 1). Among the collected data, there was only one sample of overmature forest (≥61 years) that did not meet the requirements of statistical analysis and was not considered.
Table
1.
Data classification structure of species richness of understory plants and regeneration density.
Thinning intensity (%)
Recovery time (year)
Age group (year)
HI
Thinning frequency
Species richness of understory plants
WT (≤30)
1–3
Young (≤20)
<30
Single thinning
MT (30–55)
4–6
Half-mature (21–30)
≥30
Multiple thinning
ST (>55)
7–13
Near-mature (31–40)
≥14
Mature (41–60)
Regeneration density
WT (≤30)
1–5
Half-mature (21–30)
<30
Single thinning
MT (30–50)
6–10
Near-mature (31–40)
≥30
Multiple thinning
ST (>50)
≥11
Mature (41–60)
Note: WT, MT and ST represented weak thinning, moderate thinning and strong thinning, respectively. The age groups were identified by “Regulations for age-class and age-group division of the main tree-species” (Regulatory document from the Chinese government, 2017, ICS 65.020 B60 LY, LY/T 2908-2017).
Most studies selected in this study showed only an average value of the species richness of understory plants and the density of regenerating Chinese pine seedlings and saplings, and the corresponding standard deviations or standard errors were not reported. Therefore, we used an unweighted meta-analysis to better identify the laws and characteristics of these collected data by incorporating all screened studies. The response ratio (RR) used in this study refers to the relative change in species richness of understory plants (or the density of regenerating Chinese pine seedlings and saplings) between the thinning treatment (Xe) and control treatment (Xc). The formula is defined as follows:
RR=(Xe/Xc)−1
The 95% confidence interval (CI) was estimated using the following equation:
95%CI=1.96√VR/N
where VR and N denote the variance and the number of RR for the species richness of understory plants or the density of regenerating Chinese pine seedlings and saplings, respectively. If the 95% CI does not overlap 0% change, the observed response ratio is considered significantly different from 0% change (Deng and Shangguan, 2017).
Before the meta-analysis, values outside 3 times the standard deviations of the mean RR were identified as outliers and eliminated from the analysis (Li et al., 2020). Then, the Shapiro‒Wilk test and Levene's test were used to examine the normality and homogeneity of the variances, respectively. Log(x+1) transformation was performed on the nonnormally distributed data, and the transformed data basically conformed to a normal distribution (Fig. S1). To test whether there was a significant difference in the RR of the species richness of understory plants and the density of regenerating Chinese pine seedlings and saplings between different levels, ANOVA and Duncan's multiple comparison were used to test each categorical variable, including thinning intensity, recovery time, thinning frequency, age groups, and the HI. In addition, the randomForest and Boruta packages (Kursa and Rudnicki, 2010) were used to rank the importance of factors that affect the species richness of understory plants and the density of regenerating Chinese pine seedlings and saplings, respectively. The above analysis of understory plant richness was carried out based on all understory plants (total, including shrubs and herbs), shrubs and herbs. Statistical analysis was conducted in R software (version 4.0.4).
3.
Results
3.1
Thinning intensities
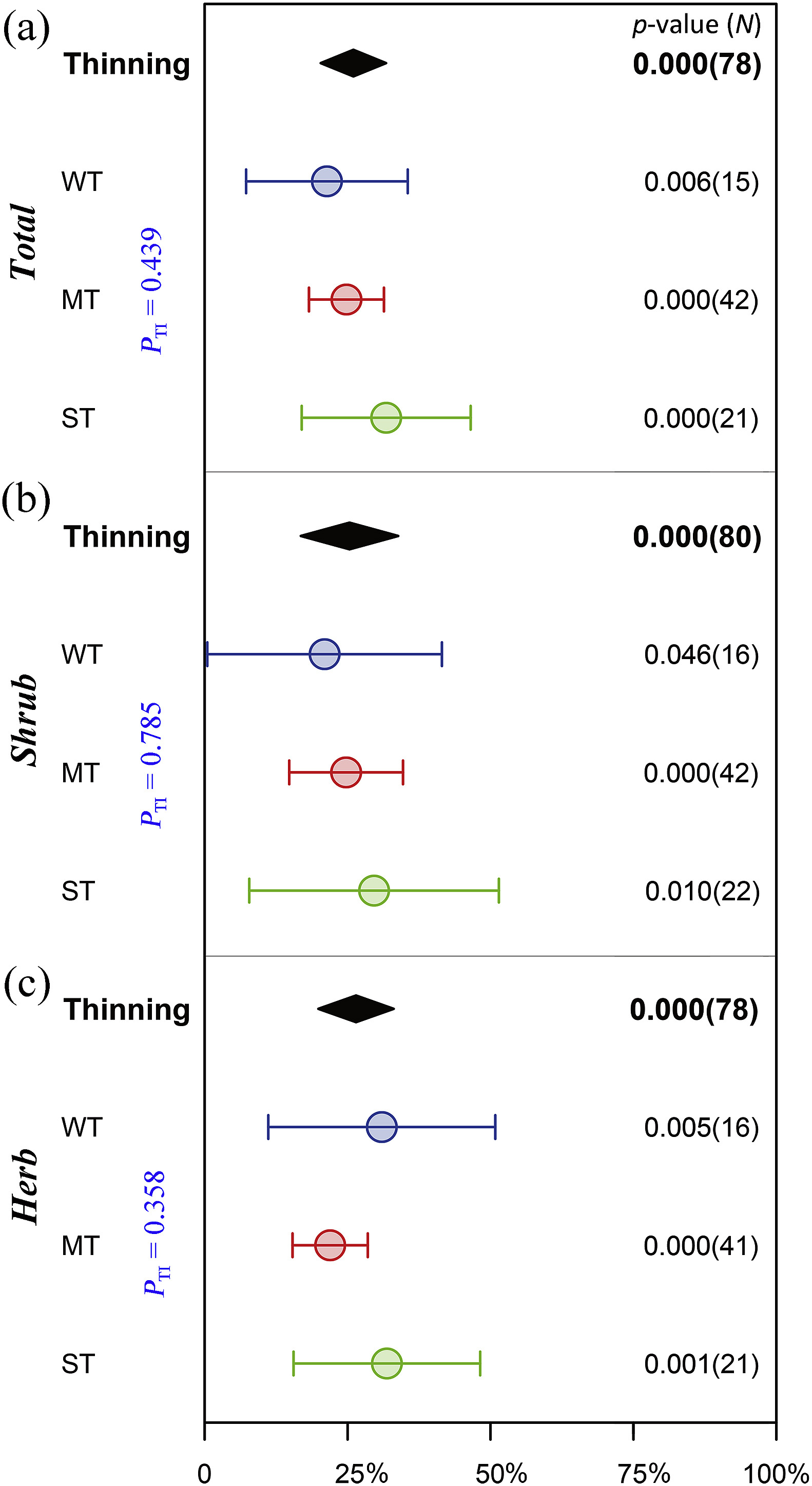
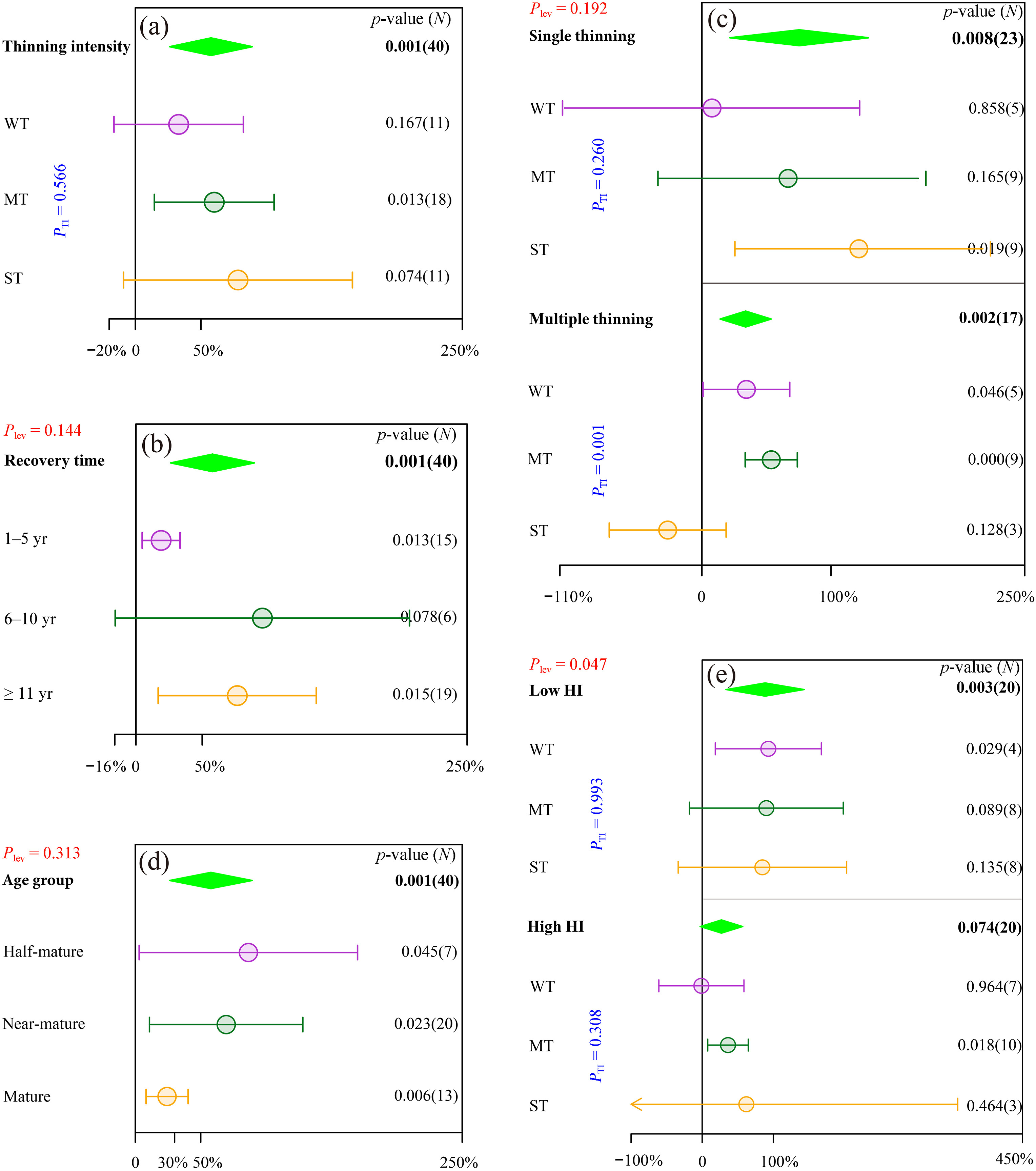
Compared with the control treatment, thinning significantly increased the species richness of shrubs and herbs by an average of 25.3% (p < 0.001) and 26.5% (p < 0.001), respectively. Moreover, different thinning intensities significantly promoted the species richness of the understory shrubs (Fig. 2b) and herbs (Fig. 2c). However, the responses of the species richness of shrubs (PTI = 0.785) and herbs (PTI = 0.358) to thinning intensity were not significant. For the density of regenerating Chinese pine seedlings and saplings, only moderate thinning had a notably positive effect (60.2%, p = 0.013, Fig. 3a).
Figure
2.
Effects of different thinning intensities on the species richness of understory plants. Dots with error bars denote the mean response ratio ±95% CI. The zero line is the reference line with a response ratio of 0. If the 95% CI overlaps with the 0 line, the effect is not significant; otherwise, it means that there is a significant effect. PTI represents the significance of the response ratio between different thinning intensities, and the center and two ends of the solid diamond represent the mean response ratio ±95% CI after combining the effects of different thinning intensities in each category. p-value (N) indicates the significance between the response ratio and the 0 line, and the sample size (the number of response ratio) for statistical analysis.
Figure
3.
Effects of thinning on the density of regenerating seedlings and saplings in Chinese pine plantations. (a) different thinning intensities; (b) different recovery time after thinning; (c) different thinning frequencies; (d) different age groups; (e) different HIs. The arrow in “(e)” indicates that the full confidence intervals of the response ratio are not shown. Plev represents the significance of response ratios between different levels of categorical variables. Please refer to Fig. 2 for other graphical descriptions.
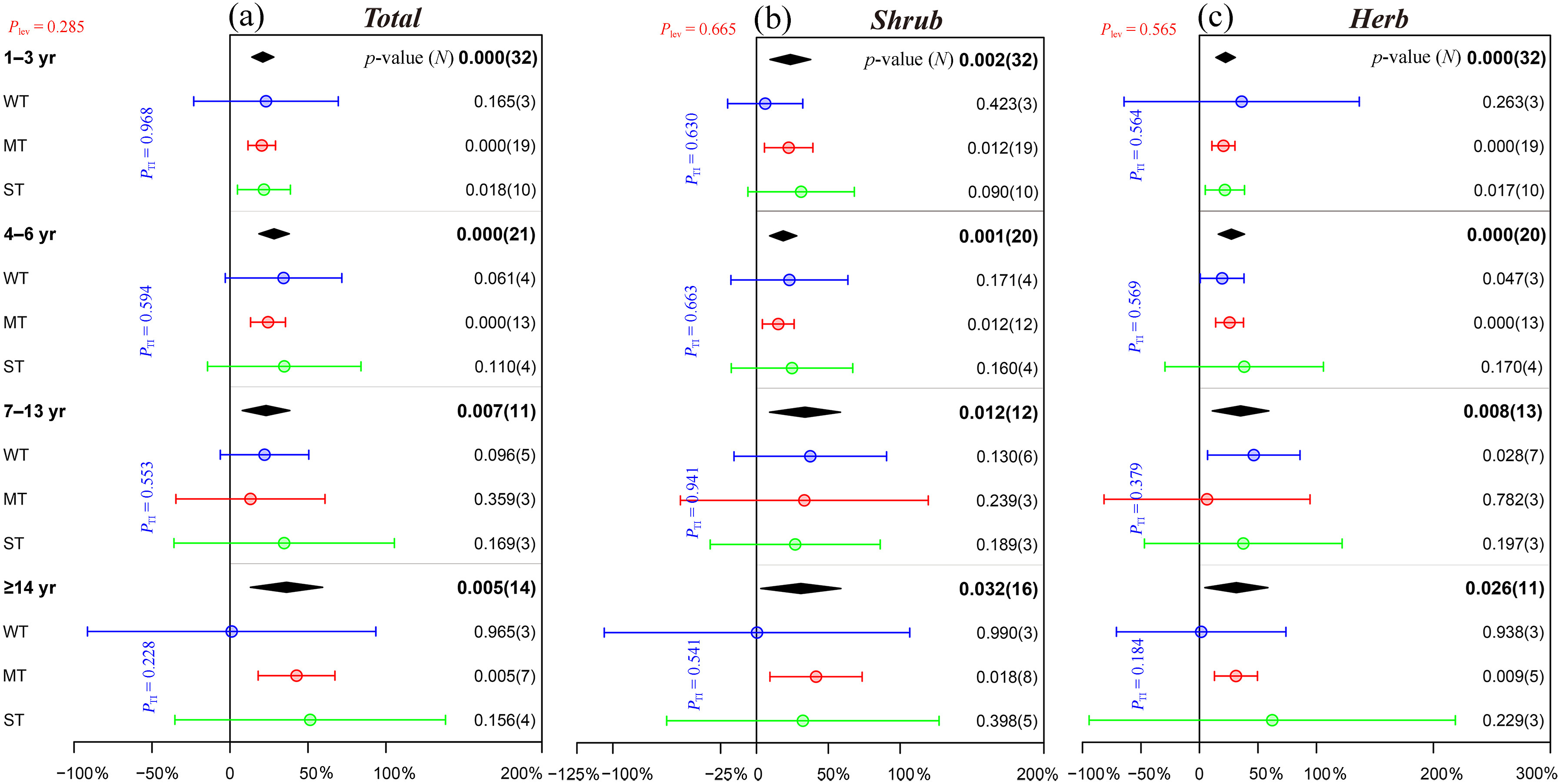
At recovery time of 1–3, 4–6, 7–13 and ≥14 years after thinning, the species richness of the understory plants was significantly increased by an average of 21.1%, 28.3%, 23.1% and 36.3%, respectively, compared to the control treatment (Fig. 4a). For understory shrubs, only moderate thinning significantly promoted species richness at different recovery time except for 7–13 years (Fig. 4b). Similar to the response of shrubs to thinning, moderate thinning apparently increased the species richness of herbs. Interestingly, strong thinning significantly increased the herb richness only in the early stage (1–3 years), while the significant effect of weak thinning was only shown after 4–13 years (Fig. 4c). For the density of regenerating Chinese pine seedlings and saplings, these notable effects occurred only at 1–5 years (19.0%, p = 0.013) and ≥11 years (76.5%, p = 0.015), although thinning significantly increased the density of regenerating Chinese pine seedlings and saplings by an average of 57.8% (p = 0.001, Fig. 3b).
Figure
4.
Effects of different recovery time after thinning on the species richness of understory plants. Please refer to Fig. 2 for graphical descriptions.
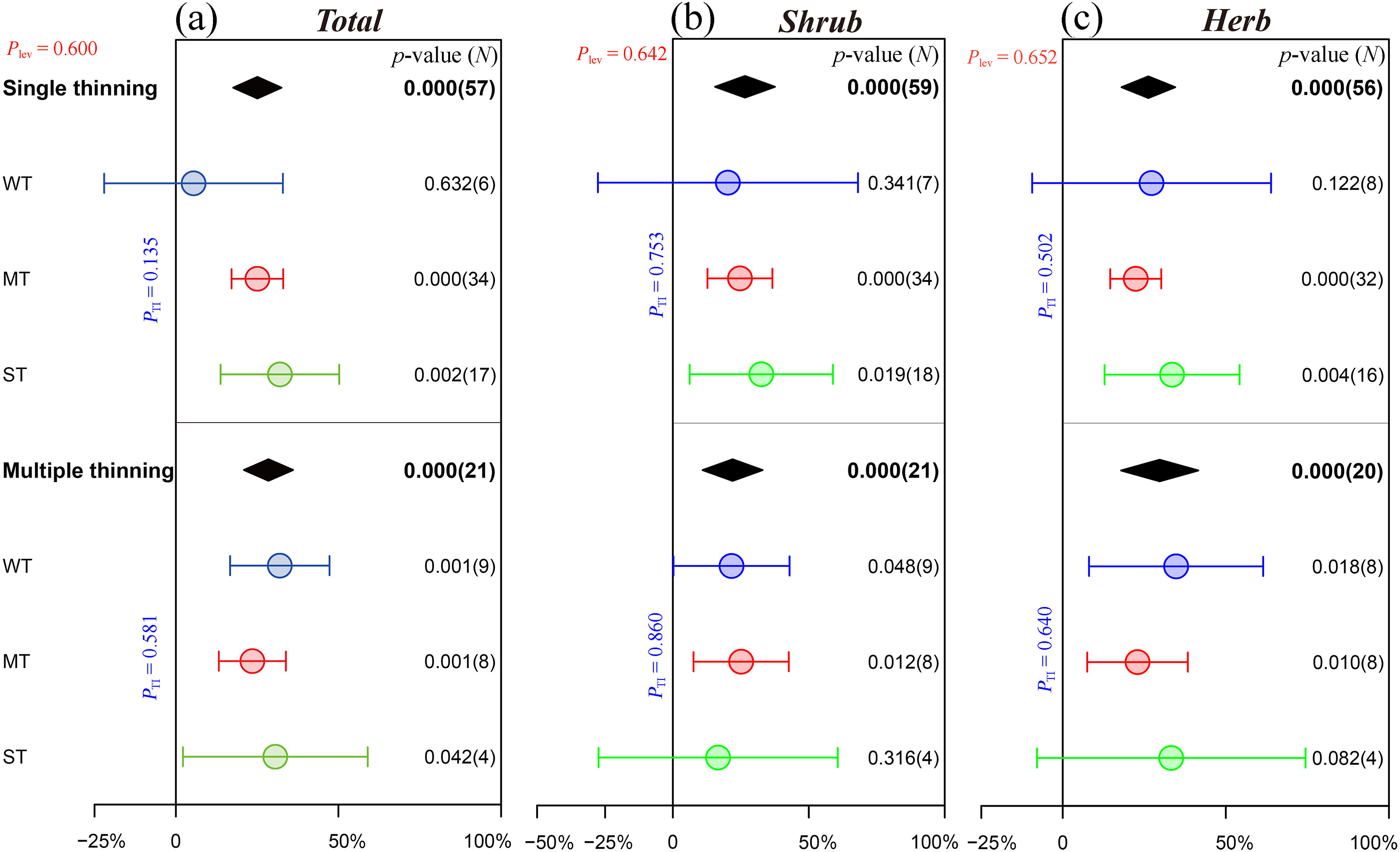
For thinning frequency, both single thinning (25.1%, p < 0.001) and multiple thinning (28.5%, p < 0.001) significantly promoted the species richness of understory plants, although there was no significant difference between them (Plev = 0.600). Specifically, except for weak thinning in single thinning and strong thinning in multiple thinning, all other thinning intensities showed an obvious promoting effect on the species richness of shrubs and herbs (Fig. 5). Similarly, single thinning (75.4%, p = 0.008) and multiple thinning (33.9%, p = 0.002) significantly increased the density of regenerating Chinese pine seedlings and saplings, and the difference between them was similar to that of the species richness, which was not significant (Plev = 0.192, Fig. 3c). Specifically, in the stands with single thinning, only strong thinning presented a pronounced increase in the density of regenerating Chinese pine seedlings and saplings (122.6%, p = 0.019). In contrast to single thinning, the effects of strong thinning on the density of regenerating Chinese pine seedlings and saplings in multiple thinning stands were 26.5% less than those of the control treatment, while weak thinning and moderate thinning significantly increased the density of regenerating Chinese pine seedlings and saplings by an average of 34.5% (p = 0.046) and 53.7% (p < 0.001), respectively (Fig. 3c).
Figure
5.
Effects of different thinning frequencies on the species richness of understory plants. Please refer to Fig. 2 for graphical descriptions.
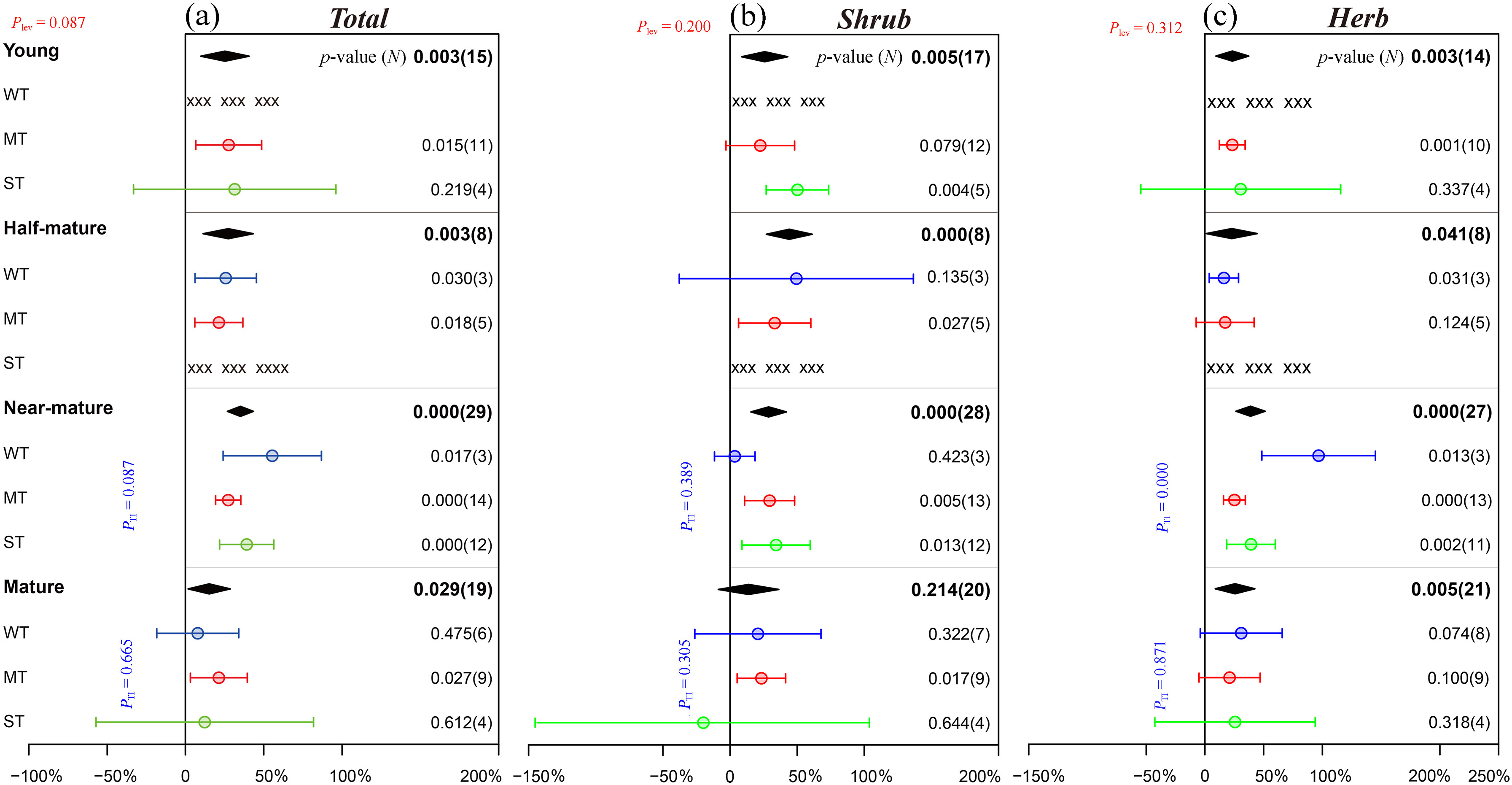
In the different age groups of Chinese pine plantations, the implementation of thinning apparently improved the species richness of understory plants. Although there was no significant difference among the different age groups (Plev = 0.087), the response of the species richness of understory plants to thinning showed a trend of first increasing and then decreasing with increasing stand age and reached a maximum (35.1%) in the near-mature forest. For the thinning intensity in different age groups, in particular, moderate thinning had a positive effect on the species richness of understory plants regardless of the forest developmental stage. Weak thinning significantly promoted the species richness of understory plants in the half-mature (p = 0.030) and near-mature forests (p = 0.017), while strong thinning had no positive effects in the young (p = 0.219) and mature forests (p = 0.612) (Fig. 6a).
Figure
6.
Effects of thinning on the species richness of understory plants in different age groups of Chinese pine plantations. “xxx” means that the number of data samples is less than 3 and without statistical analysis. Please refer to Fig. 2 for graphical descriptions.
In addition to mature forests (p = 0.214), thinning actively promoted the increase in shrub species richness in young, half-mature and near-mature forests by an average of 25.8% (p = 0.005), 44.1% (p < 0.001) and 28.7% (p < 0.001), respectively. In all age groups, the promotion effect of moderate thinning on the species richness of shrubs was significant except in young forests (p = 0.079), while the exhibited effect of weak thinning was not obvious. Strong thinning had a significant positive effect on shrub richness in young (p = 0.004) and near-mature forests (p = 0.013), but weak inhibition was observed in mature forests (−19.9%, p = 0.644) (Fig. 6b). Similarly, different thinning intensities significantly promoted the species richness of herbs in young (p = 0.003), half-mature (p = 0.041), near-mature (p < 0.001) and mature forests (p = 0.005), although the difference among them was not significant (Plev = 0.312, Fig. 6c).
With respect to the density of regenerating Chinese pine seedlings and saplings in different age groups, thinning had a significantly positive effect, with an average RR of 57.8% (p = 0.001). However, the difference among age groups was not significant (Plev = 0.313). Interestingly, with the succession of forest stands, the promoting effect of thinning on the density of regenerating Chinese pine seedlings and saplings decreased gradually, and the largest thinning effect appeared in the half-mature forests (86.5%, p = 0.045, Fig. 3d).
3.5
Humidity indexes
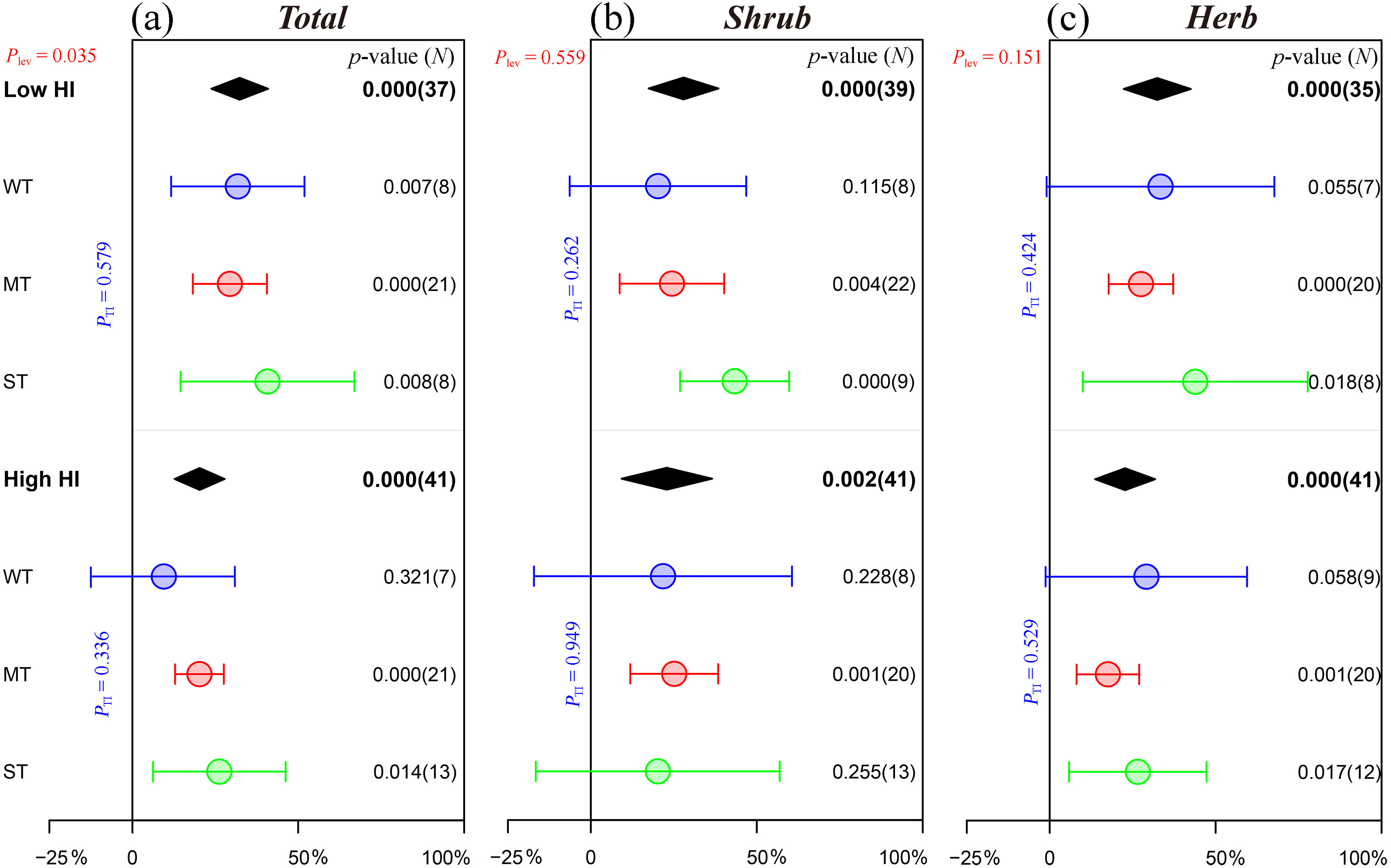
After thinning in areas with different HIs, it was found that the species richness of understory plants was significantly improved in Chinese pine plantations, and the effects of thinning in low-humidity areas (HI < 30) were significantly better than those in high-humidity areas (HI ≥ 30, Plev = 0.035). For the different thinning intensities in areas with different HIs, the thinning performances were different. In particular, only moderate and strong thinning in low-humidity areas and moderate thinning in high-humidity areas significantly increased the species richness of shrubs and herbs (Fig. 7). Similar to the species richness of understory plants, the promoting effects of thinning on the density of regenerating Chinese pine seedlings and saplings in low-humidity areas were greater than those in high-humidity areas (Plev = 0.047) and had a positive effect only in low-humidity areas (p = 0.003). In low-humidity and high-humidity areas, only weak thinning (p = 0.029) and moderate thinning (p = 0.018) significantly increased the density of regenerating Chinese pine seedlings and saplings, respectively (Fig. 3e).
Figure
7.
Effects of thinning on the species richness of understory plants of Chinese pine plantations in different climatic conditions. Please refer to Fig. 2 for graphical descriptions.
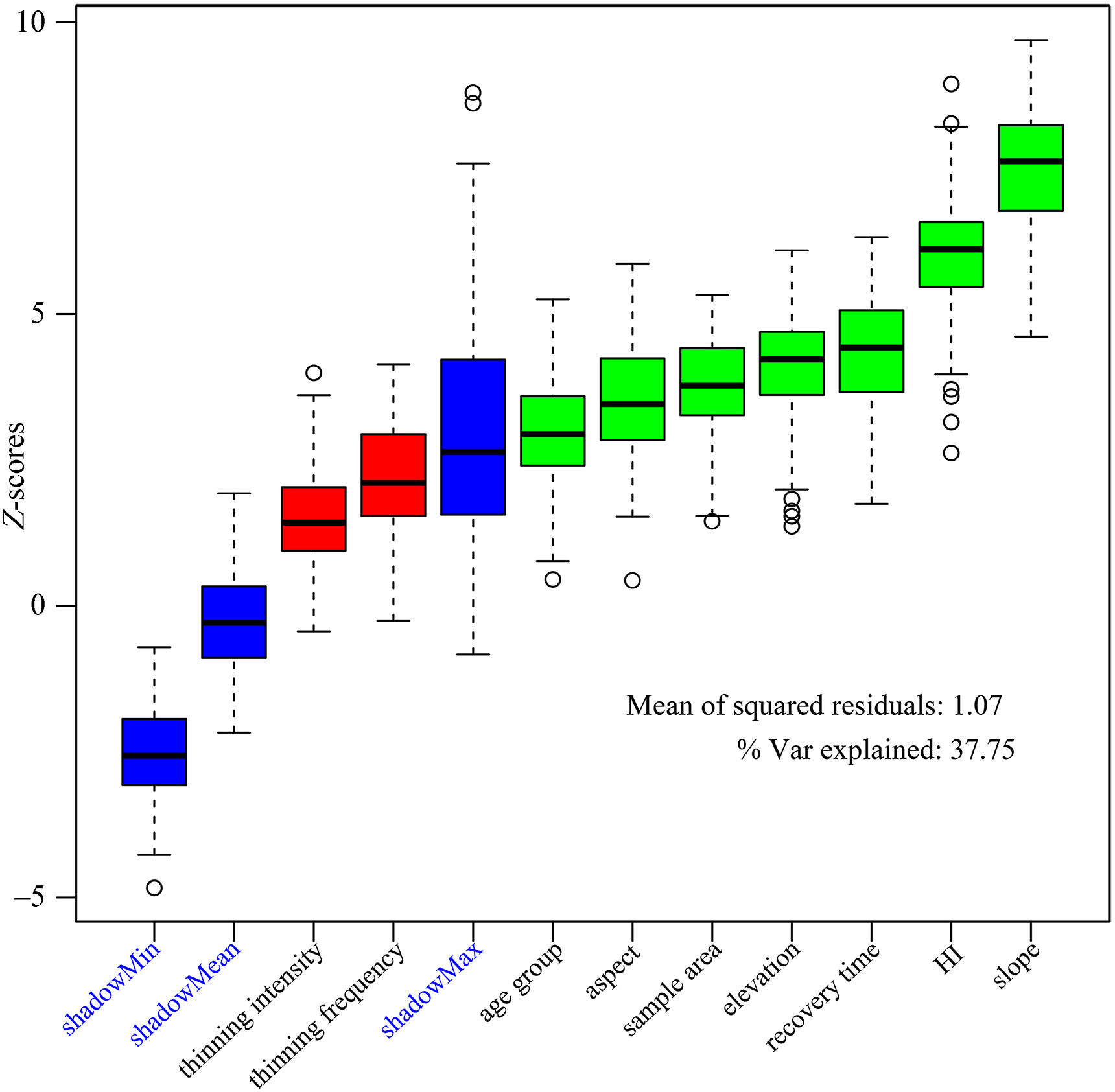
All screened environmental factors were analyzed, and age group, planting density, recovery time, elevation and MAT were the important factors affecting the species richness of all understory plants in the Chinese pine plantation. Among the top five important factors affecting the species richness of shrubs and herbs, there were 4 common environmental factors, including planting density, elevation, recovery time and age group (Table 2). Nonetheless, these vital factors had different effects on species richness. For example, planting density had the greatest impacts on shrubs, followed by slope, while the largest contributor to herbs was age group and then elevation. In addition, thinning frequency had a greater impact on herbs than did the other 7 factors (Table 2). The above analyses indicated that shrubs and herbs had different sensitivities to environmental factors. For the density of regenerating Chinese pine seedlings and saplings, the environmental factors involved in this study explained 37.75% of their variation, and the order of importance of environmental factors was as follows: slope (7.53%) > HI (5.98%) > recovery time (4.35%) > elevation (4.07%) > sample area (3.75%) > aspect (3.49%) > age group (2.91%) (Fig. 8).
Table
2.
Factors affecting the species richness of understory plants in Chinese pine plantation.
Factors
%IncMSE
Total
Shrub
Herb
MAT
6.64
3.20
6.64
MAP
3.31
3.26
3.25
Aspect
2.52
1.49
3.49
Slope
4.86
8.59
6.36
Elevation
8.93
6.07
10.83
Planting density
11.42
11.85
9.85
Retention density
3.01
0.58
7.24
Age group
11.68
3.98
14.63
Thinning frequency
4.43
3.44
7.33
Thinning intensity
−1.00
−2.76
−0.10
Recovery time
9.88
4.31
9.68
Sample area
3.63
0.00
Note: MAT = mean annual temperature; MAP = mean annual precipitation. These statistics were evaluated by the randomForest function in randomForest package of R.
Figure
8.
Factors affecting the density of regenerating seedlings and saplings in Chinese pine plantations. The red boxplots represent the factors that had little impact on the density of regenerating Chinese pine seedlings and saplings and were removed. The green boxplots represent the factors that had a greater impact on the density of regenerating Chinese pine seedlings and saplings and were retained. The blue boxplots represent the minimal, average and maximum Z-scores of the shadow attributes. If the importance of an attribute is significantly lower than shadowMax, it will be considered unimportant and permanently deleted; if the importance of an attribute is significantly higher than shadowMax, it will be considered important. The analysis was performed by the Boruta function in the Boruta package of R software. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
In addition to the categorical variables described above, we conducted a relationship analysis between other factors and the RR. The results suggested that slope had a significant effect on the species richness of shrubs (Estimate = −1.249, p = 0.002) and the density of regenerating Chinese pine seedlings and saplings (Estimate = −1.826, p = 0.007) and exhibited a notable negative correlation with them. In general, the probabilities that thinning could promote the species richness of shrubs and herbs and the density of regenerating Chinese pine seedlings and saplings were 74.70%, 87.65% and 76.74%, respectively (Table 3).
Table
3.
Classification statistics on the correlations between thinning and the species richness of understory plants and natural regeneration density.
Variable
Sample size (N)
+
–
0
Species richness of understory plants
RR-T
83
75 (90.36%)
7 (8.43%)
1 (1.20%)
RR-S
83
62 (74.70%)
12 (14.46%)
9 (10.84%)
RR-H
81
71 (87.65%)
9 (11.11%)
1 (1.23%)
Regeneration density
RR-d
43
33 (76.74%)
10 (23.26%)
0
Note: The symbols +, −, and 0 represent positive relationships, negative relationships, and no relationship, respectively. N represents the number of RR. RR-T, RR-S, RR-H represent the response ratio of the species richness of all understory plants, shrubs and herbs. RR-d represents the response ratio of regeneration density.
Our study mainly analyzed and quantified the effects of thinning on the species richness of understory plants in Chinese pine plantations and found that different thinning intensities could significantly improve the species richness of understory plants (Fig. 2), which was consistent with the results of some previous studies (Chen and Cao, 2014; Willms et al., 2017; Dang et al., 2018). This may be attributed to the fact that ecological space and light in the forest increase after thinning (Hale, 2003). In turn, these changed environments accelerate the decomposition of litter and increase the availability of nutrients and water in the soil (Trentini et al., 2017; Zhang et al., 2018). Ultimately, the heterogeneity of the understory microenvironment will be enriched; thus, suitable conditions will be created to meet the coexistence of more plant species. In addition, the high planting density (generally >2500 stems·ha−1) of Chinese pine plantations may lead to the positive effect of thinning increases with the strengthening of thinning intensity before the forest stands reach the most reasonable density (Chen and Cao, 2014).
We found that with the increase in thinning intensity, the species richness of the understory plants and the density of regenerating Chinese pine seedlings and saplings increased, especially moderate thinning, which caused a significant increase of 60.2% in the density of regenerating Chinese pine seedlings and saplings (Fig. 3a). This finding was similar to that of Olson et al. (2014), who found that the rate of recruitment increased as the thinning intensity increased, thus stimulating the occurrence of understory regeneration similar to that of a shelterwood establishment cut, resulting in an increase in the regeneration density. However, we found that the effects of weak thinning and strong thinning on the density of regenerating Chinese pine seedlings and saplings were not significantly different. This phenomenon may be better explained by the intermediate disturbance hypothesis, which states that the highest diversity is maintained at an intermediate scale of disturbance (Connell, 1978). It is possible that the excessive thinning intensity exacerbated the evaporation of soil water and finally caused drought stress, limiting the early establishment and growth of regenerated seedlings of Chinese pine plantations in northern China (Lei et al., 2018; Deng et al., 2020). In addition, the low intensity of thinning could not effectively improve the environmental conditions of depressed forest stands or alleviate the fierce competition for limited resources.
4.2
Influence of recovery time
Recovery time is an indispensable factor in evaluating the responses of forest restoration to disturbance (Crouzeilles et al., 2016), but there are still controversies in some previous studies about the responses of the species richness of understory plants to thinning during different recovery time. For example, Li et al. (2020) found that the positive impacts of thinning on the species richness of understory shrubs and herbs can last for a long time, and species richness reached a maximum in stands with a recovery time longer than 20 years after thinning. Similarly, our study found that the species richness of understory plants in the Chinese pine plantation was dramatically improved at different recovery time after thinning (Fig. 4), which indirectly indicated that the positive promotion of thinning on understory plants had a long-term effect. In contrast, some studies (Ma et al., 2007; Chen et al., 2014; Zhou et al., 2016) identified that thinning has no long-term effect on the improvement of the species richness of understory plants in Chinese pine plantations. They maintained the opinion that with the gradual closure of the forest stand, the environmental heterogeneity would gradually disappear, thereby resulting in the species and number of understory plants being lower than those in the early period of thinning. The differences in the thinning effects in these studies may be caused by their different site conditions, stand characteristics and research scales.
In addition, we found that the recovery time of 1–5 and ≥11 years after thinning both actively promoted the regeneration density of Chinese pine, while the recovery time of 6–10 years had no significant effect on the density of regenerating Chinese pine seedlings and saplings (Fig. 3b). This result may be because the environmental conditions under a closed canopy improved during the initial period of thinning, which increased the available nutrients in the woodland (del Campo et al., 2019), thereby promoting the germination and establishment of Chinese pine seedlings. Nevertheless, with the development of forest succession, the favorable ecological niche provided by thinning in the stand may also change (Huang et al., 2014), and the competition among regenerated seedlings and between them and understory shrubs and herbs gradually increased, leading to a decline in the density of regenerated seedlings. Moreover, thinning improves the microenvironment in the forest while promoting the expansion and healthy growth of the crown width of the retained mature tree in the stands, thereby gradually enhancing the yield and quality of seeds, which may be one of the reasons for the significant increase in the density of regenerated seedlings at a recovery time of ≥11 years after thinning (Flathers et al., 2016; Liang and Wei, 2020).
4.3
Influence of thinning frequency
In this study, both single thinning and multiple thinning significantly increased the species richness of understory plants and the regeneration density of Chinese pine plantations. However, for different thinning intensities, only strong thinning in single thinning and weak and moderate thinning in multiple thinning significantly promoted the species richness of the understory plants (Figs. 5 and 3c). This finding was similar to the conclusion of Goodwin et al. (2018), who found that thinning reduced the thickness of litter and increased the availability of bare ground during the 15 years after thinning, but these positive effects were short-lived in the absence of a secondary disturbance. Therefore, for the stands with single thinning, a stronger thinning intensity was required to expand the effective space and improve the light environment for plant growth. For the stands that were thinned many times, a lighter thinning intensity was required to avoid sudden and drastic changes in the microenvironment, which are not conducive to the survival of understory plants. Therefore, we suggest that for stands that have experienced multiple thinning in their management history, a gentler thinning intensity should be used to prevent the positive effect of thinning from decreasing or disappearing when thinning is carried out again.
4.4
Influence of age group
We found that thinning could significantly increase the species richness of understory plants and the density of regenerating seedlings and saplings in different age groups of Chinese pine plantations, and the positive effect began to present a decreasing trend from the half-mature forests (Figs. 6 and 3d). We postulated that the amount of available resources and their heterogeneity were one of the main reasons for the occurrence of this trend because the effects of the former mainly occurred in the early stage of the stands, while the impacts of the latter primarily appeared in the later period of the stands (Bartels and Chen, 2010). Alternatively, another major reason for this trend may be attributed to the gradual decrease in effective living space with stand succession. In the early stages of forest succession, understory plants had higher productivity and higher spatial heterogeneity benefited from the weak dominance of canopy trees (Swanson et al., 2011). With the growth and development of canopy trees, the gradual closure of the canopy space led to the reduction of light in the forest (Azeria et al., 2011) and reduced the available resources and limited living space, resulting in fierce intraspecific and interspecific competition between regenerated seedlings and other understory plants, which weakened the positive promotion of thinning on the species richness of understory plants and the regeneration density to a certain extent. Based on our study, we suggest that in the middle stage of stand succession (half-mature forest), thinning should be carried out in a timely manner to maximize the potential promotion of thinning on understory plant diversity and natural regeneration, thus maximizing the ecological service function of soil and water conservation and realizing the continuous coverage of Chinese pine plantations.
4.5
Influence of other factors involved
On the scale of northern China, we found that there were certain differences in key factors affecting the species richness of understory plants and the density of regenerating Chinese pine seedlings and saplings after thinning. For example, stand factors such as age group, planting density, and recovery time were important factors affecting the former, while the slope, HI and recovery time were vital factors affecting the latter (Table 2, Fig. 8). This finding provides some insights for future forest management: targeted tending measures should be adopted according to the different objectives of forest cultivation. For example, if plantation stands are focused on ecological restoration and protection, such as soil and water conservation, a reasonable interval of recovery time after thinning should be controlled for different forest development stages to achieve the purpose of having a profound impact on the species diversity of understory plants. If the purpose is to achieve sustainable development of forest stands by utilizing the natural regeneration capacity of plantations, we should first consider the decisive influence of climatic conditions (e.g., HI) and site conditions (e.g., slope) on the density of regenerating Chinese pine seedlings and saplings. In other words, the climatic conditions and slope in which the stands are located may directly affect the success of natural regeneration of Chinese pine plantations. In addition, it is necessary to consider the long-term (≥11 years) responses of the density of regenerated seedlings to thinning, which suggests that some active measures, such as soil loosening, moderate clearing and gap expansion, should be supplemented during this period to promote the occurrence and survival of natural regeneration.
As discussed above, planting density, slope and HI had a greater impact than other factors on the species richness of understory plants and the density of regenerating Chinese pine seedlings and saplings. Specifically, the planting density and thinning intensity could directly control the stand density, which in turn determined the openness of the canopy and indirectly had an impact on the species richness and composition of understory plants. When the stand density is too low, the species differentiation of the understory vegetation is intense, and the population competition will be fierce, while when the stand density is too large, a large number of canopy trees occupy too much living space and resources, and the inhibitory effect on the understory plants will be enhanced (Kang et al., 2009). Slope, aspect and altitude are important site factors that affect the distribution and growth of understory plants. Their impacts on richness and natural regeneration were mainly caused by the redistribution of ecological factors such as light, heat, water and soil nutrients (Takahashi, 2010; Liu and Wang, 2013).
In addition, the responses of the species richness of understory plants and the density of regenerating Chinese pine seedlings and saplings to thinning in low-HI areas were significantly higher than those in high-HI areas (Figs. 3e and 7). This result was because the native species pools in arid areas were relatively small compared with those in humid areas, resulting in a more sensitive response of the species richness of understory plants to thinning in arid areas (Li et al., 2020). Ultimately, the increase in the species richness of understory plants in low-HI areas was higher than that in high-HI areas after thinning. In addition, after thinning in low-HI areas, the competition of plants for limited water resources was reduced, and the water infiltration of soil and surface runoff were increased, which created relatively better conditions for the recruitment, establishment and settlement of understory plants (Sohn et al., 2016; del Campo et al., 2019). We also found that the explanatory power of thinning intensity and thinning frequency on the species richness of understory plants and the density of regenerating Chinese pine seedlings and saplings was much smaller than that of other factors (Table 2, Fig. 8), which may be due to the impact of anthropogenic interference (e.g., thinning intensity) on the dynamic changes in understory plants being relatively weak compared with large-scale environmental factors.
5.
Conclusions
We found that the species richness of understory plants exhibited a significantly positive response to different thinning intensities and increased with increasing thinning intensities. However, only moderate thinning significantly promoted the density of regenerating Chinese pine seedlings and saplings. Interestingly, these promoting effects on understory plants were long-term, and the species richness of understory plants after 14 years of thinning was largest, while the density of regenerating Chinese pine seedlings and saplings reached a maximum at recovery time ≥11 years. Among different age groups, the maximum thinning responses of shrub species richness and regeneration density both occurred in the half-mature Chinese pine plantations, which were 44.1% and 86.5% higher than those in the control stands, respectively.
In general, age group, planting density, and recovery time were important factors influencing the species richness of understory plants, while the slope, HI, and recovery time had the greatest impacts on the density of regenerating Chinese pine seedlings and saplings. This result implies that the responses of the species richness of understory plants and the density of regenerating Chinese pine seedlings and saplings to these key factors were different after thinning. Therefore, we suggest that if plantation forests focus on ecological restoration, such as soil and water conservation, a reasonable restoration interval after thinning should be controlled for different forest development stages to maximize the species diversity of understory plants. If the purpose of cultivation is to achieve sustainable development by utilizing the natural regeneration capacity of plantations, we should first consider the decisive influence of climatic conditions (such as the HI) and stand conditions (such as the slope) on the density of regenerated seedlings. Then, some positive measures, such as soil loosening, moderate clearing and gap expansion, should be supplemented after thinning to promote the occurrence and survival of natural regeneration.
Author contributions
Haijiao Yang, Chao Pan, Zhibin Wang, and Dihai Wang conceived the study and planned the work. Yun Wu and Shiqi Qing prepared the dataset. Haijiao Yang, Chao Pan and Zhibin Wang, wrote the majority of the manuscript. All authors read and approved the final manuscript.
Availability of data and materials
The dataset analyzed during the current study is shown in tables, figures and supplementary materials, further inquiries can be directed to the corresponding author.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Declaration of competing interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Aavik, T., Püssa, K., Roosaluste, E., Moora, M., 2009. Vegetation change in boreonemoral forest during succession-trends in species composition, richness and differentiation diversity. Ann. Bot. Fenn. 46 (4), 326-335. .
[2]
Ali, A., Lin, S.L., He, J.K., Kong, F.M., Yu, J.H., Jiang, H.S., 2019. Climate and soils determine aboveground biomass indirectly via species diversity and stand structural complexity in tropical forests. For. Ecol. Manag. 432, 823-831. .
[3]
An, Y., Ding, G.D., Liang, J.H., Gao, G.L., He, Y., Wei, B., Bao, Y.F., Bao, B., 2012. Effects of thinning on the growth and the development of undergrowth of Pinus tabulaeformis plantation in Rocky Mountain area of north China. Res. Soil Water Conserv. 19, 86–90 (in Chinese with English abstract).
[4]
Aun, K., Kukumägi, M., Varik, M., Becker, H., Aosaar, J., Uri, M., Morozov, G., Buht, M., Uri, V., 2021. Short-term effect of thinning on the carbon budget of young and middle-aged Scots pine (Pinus sylvestris L.) stands. For. Ecol. Manag. 492, 119241. .
[5]
Azeria, E.T., Bouchard, M., Pothier, D., Fortin, D., Hébert, C., 2011. Using biodiversity deconstruction to disentangle assembly and diversity dynamics of understory plants along post-fire succession in boreal forest. Global Ecol. Biogeogr. 20 (1), 119-133. .
[6]
Bartels, S.F., Chen, H.Y.H., 2010. Is understory plant species diversity driven by resource quantity or resource heterogeneity? Ecology 91 (7), 1931-1938. .
[7]
Chen, Y.M., Cao, Y., 2014. Response of tree regeneration and understory plant species diversity to stand density in mature Pinus tabulaeformis plantations in the hilly area of the Loess Plateau, China. Ecol. Eng. 73, 238-245. .
[8]
Connell, J.H., 1978. Diversity in tropical rain forests and coral reefs. Science 199 (4335), 1302-1310. .
[9]
Crouzeilles, R., Curran, M., Ferreira, M.S., Lindenmayer, D.B., Grelle, C.E.V., Benayas, J.M.R., 2016. A global meta-analysis on the ecological drivers of forest restoration success. Nat. Commun. 7, 11666.
[10]
Dang, P., Gao, Y., Liu, J.L., Yu, S.C., Zhao, Z., 2018. Effects of thinning intensity on understory vegetation and soil microbial communities of a mature Chinese pine plantation in the Loess Plateau. Sci. Total Environ. 630, 171-180. .
[11]
del Campo, A.D., González-Sanchis, M., Molina, A.J., García-Prats, A., Ceacero, C.J., Bautista, I., 2019. Effectiveness of water-oriented thinning in two semiarid forests: the redistribution of increased net rainfall into soil water, drainage and runoff. For. Ecol. Manag. 438, 163-175. .
[12]
Deng, C., Zhang, S.G., Lu, Y.C., Froese, R.E., Xu, X.J., Zeng, J., Ming, A.G., Liu, X.Z., Xie, Y.S., Li, Q.F., 2020. Thinning effects on forest evolution in Masson pine (Pinus massoniana Lamb.) conversion from pure plantations into mixed forests. For. Ecol. Manag. 477, 118503. .
[13]
Deng, L., Shangguan, Z.P., 2017. Afforestation drives soil carbon and nitrogen changes in China. Land Degrad. Dev. 28 (1), 151-165. .
[14]
Dupuy, J.M., Chazdon, R.L., 2008. Interacting effects of canopy gap, understory vegetation and leaf litter on tree seedling recruitment and composition in tropical secondary forests. For. Ecol. Manag. 255 (11), 3716-3725. .
[15]
FAO, 2020. Global Forest Resources Assessment 2020 (FRA 2020). FRA 2020 Working Paper. UN Food and Agriculture Organization, Rome. . (Accessed 15 September 2022).
Gao, Y.C., Zhang, W.H., He, J.F., Wang, J.T., 2013. Effects of thinning intensity on Pinus tabulaeformis plantation in Huanglong Mountain, Northwest China: a comprehensive evaluation. Chin. J. Appl. Ecol. 24 (5), 1313–1319. (in Chinese with English abstract).
Goodwin, M.J., North, M.P., Zald, H.S.J., Hurteau, M.D., 2018. The 15-year post-treatment response of a mixed-conifer understory plant community to thinning and burning treatments. For. Ecol. Manag. 429, 617-624. .
[20]
Han, W.J., Cao, X.P., Zhang, W.H., 2014. Effect of ground cover on early regeneration of Pinus tabulaeformis plantation. Sci. Silvae Sin. 50, 49-54. (in Chinese with English abstract).
[21]
Hale, S.E., 2003. The effect of thinning intensity on the below canopy light environment in a Sitka spruce plantation. For. Ecol. Manag. 179 (1–3), 341-349. .
[22]
Huang, P.S., Lin, H.C., Lin, C.P., Tso, I.M., 2014. The effect of thinning on ground spider diversity and microenvironmental factors of a subtropical spruce plantation forest in East Asia. Eur. J. For. Res. 133 (5), 919-930. .
[23]
Ito, S., Ishigami, S., Mizoue, N., Buckley, G.P., 2006. Maintaining plant species composition and diversity of understory vegetation under strip-clearcutting forestry in conifer plantations in Kyushu, southern Japan. For. Ecol. Manag. 231, 234-241. .
[24]
Jiménez, M.N., Spotswood, E.N., Cañadas, E.M., Navarro, F.B., 2015. Stand management to reduce fire risk promotes understorey plant diversity and biomass in a semi-arid Pinus halepensis plantation. Appl. Veg. Sci. 18 (3), 467-480. .
[25]
Kang, B., Liu, S.R., Cai, D.X., Lu, L.H., 2009. Effects of Pinus massoniana plantation stand density on understory vegetation and soil properties. Chin. J. Appl. Ecol. 20 (10), 2323–2331 (in Chinese with English abstract)
[26]
Kursa, M.B., Rudnicki, W.R., 2010. Feature selection with Boruta package. J. Stat. Software 36 (11), 1-13.
[27]
Lei, L., Xiao, W.F., Zeng, L.X., Zhu, J.H., Huang, Z.L., Cheng, R.M., Gao, S.K., Li, M., 2018. Thinning but not understory removal increased heterotrophic respiration and total soil respiration in Pinus massoniana stands. Sci. Total Environ. 621, 1360-1369. .
[28]
Li, X.K., Li, Y.A., Zhang, J., Peng, S.Z., Chen, Y.M., Cao, Y., 2020. The effects of forest thinning on understory diversity in China: a meta-analysis. Land Degrad. Dev. 31 (10), 1225-1240. .
[29]
Liang, W.J., Wei, X., 2020. Factors promoting the natural regeneration of Larix principis-rupprechtii plantation in the Lvliang Mountains of central China. PeerJ 8, e9339. .
[30]
Liu, S.R., Yang, Y.J., Wang, H., 2018. Development strategy and management countermeasures of planted forests in China: transforming from timber-centered single objective management towards multi-purpose management for enhancing quality and benefits of ecosystem services. Acta Ecol. Sin. 38 (1), 1-10. (in Chinese with English abstract).
[31]
Liu, M.X., Wang, G., 2013. Responses of plant community diversity and soil factors to slope aspect in alpine meadow. Chin. J. Ecol. 32, 259-265.
[32]
Ma, L.Y., Li, C.Y., Wang, X.Q., Xu, X., 2007. Effects of thinning on the growth and the diversity of undergrowth of Pinus tabulaeformis plantation in Beijing mountainous areas. Sci. Silv. Sin. 43, 1-9. (in Chinese with English abstract).
[33]
Mori, A.S., Furukawa, T., Sasaki, T., 2013. Response diversity determines the resilience of ecosystems to environmental change. Biol. Rev. 88, 349-364. .
[34]
Nilsson, M.C., Wardle, D.A., 2005. Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest. Front. Ecol. Environ. 3 (8), 421-428. .
Pan, Y.D., Birdsey, R.A., Fang, J.Y., Houghton, R., Kauppi, P.E., Kurz, W.A., Phillips, O.L., Shvidenko, A., Lewis, S.L., Canadell, J.G., Ciais, P., Jackson, R.B., Pacala, S.W., McGuire, A.D., Piao, S.L., Rautiainen, A., Sitch, S., Hayes, D., 2011. A large and persistent carbon sink in the world's forests. Science 333 (6045), 988-993. .
[37]
Raymond, P., Royo, A.A., Prévost, M., Dumais, D., 2018. Assessing the single-tree and small group selection cutting system as intermediate disturbance to promote regeneration and diversity in temperate mixedwood stands. For. Ecol. Manag. 430, 21-32. .
[38]
Sabo, K.E., Sieg, C.H., Hart, S.C., Bailey, J.D., 2009. The role of disturbance severity and canopy closure on standing crop of understory plant species in ponderosa pine stands in northern Arizona, USA. For. Ecol. Manag. 257, 1656-1662. .
[39]
Seiwa, K., Negishi, Y., Eto, Y., Hishita, M., Negishi, S., Masaka, K., Suzuki, M., 2021. Effects of repeated thinning at different intensities on the recovery of hardwood species diversity in a Cryptomeria japonica plantation. J. For. Res. 26 (1), 17-25. .
[40]
Sohn, J.A., Saha, S., Bauhus, J., 2016. Potential of forest thinning to mitigate drought stress: a meta-analysis. For. Ecol. Manag. 380, 261-273. .
[41]
State Forestry Administration, 2018. National Forest Resources Statistics: the Eighth National Forest Resources Inventory. China Forestry Publishing House, Beijing.
Takahashi, K., 2010. Effects of altitude and competition on growth and mortality of the conifer Abies sachalinensis. Ecol. Res. 25 (4), 801-812. .
[44]
Trentini, C.P., Campanello, P.I., Villagra, M., Ritter, L., Ares, A., Goldstein, G., 2017. Thinning of loblolly pine plantations in subtropical Argentina: impact on microclimate and understory vegetation. For. Ecol. Manag. 384, 236-247. .
[45]
Wang, Z.B., Yang, H.J., Dong, B.Q., Zhou, M.M., Ma, L.Y., Jia, Z.K., Duan, J., 2017. Effects of canopy gap size on growth and spatial patterns of Chinese pine (Pinus tabulaeformis) regeneration. For. Ecol. Manag. 385, 46-56. .
[46]
Wang, Z.B., Yang, H.J., Wang, D.H., Zhao, Z., 2019a. Spatial distribution and growth association of regeneration in gaps of Chinese pine (Pinus tabuliformis Carr.) plantation in northern China. For. Ecol. Manag. 432, 387-399. .
[47]
Wang, Z.B., Yang, H.J., Wang, D.H., Zhao, Z., 2019b. Response of height growth of regenerating trees in a Pinus tabulaeformis Carr. plantation to different thinning intensities. For. Ecol. Manag. 444, 280-289. .
[48]
Wang, D., Olatunji, O.A., Xiao, J.L., 2019c. Thinning increased fine root production, biomass, turnover rate and understory vegetation yield in a Chinese fir plantation. For. Ecol. Manag. 440, 92-100. .
[49]
Willms, J., Bartuszevige, A., Schwilk, D.W., Kennedy, P.L., 2017. The effects of thinning and burning on understory vegetation in North America: a meta-analysis. For. Ecol. Manag. 392, 184-194. .
[50]
Wilsey, B.J., Chalcraft, D.R., Bowles, C.M., Willig, M.R., 2005. Relationships among indices suggest that richness is an incomplete surrogate for grassland biodiversity. Ecology 86 (5), 1178-1184. .
[51]
Xu, H., 1993. Chinese Pine (Pinus tabulaeformis). China Forestry Publishing House, Beijing (In Chinese)
[52]
Zhang, X.Z., Guan, D.X., Li, W.B., Sun, D., Jin, C.J., Yuan, F.H., Wang, A.Z., Wu, J.B., 2018. The effects of forest thinning on soil carbon stocks and dynamics: a meta-analysis. For. Ecol. Manag. 429, 36-43. .
[53]
Zhong, Z.K., Wang, X., Zhang, X.Y., Zhang, W., Xu, Y.D., Ren, C.J., Han, X.H., Yang, G.H., 2019. Edaphic factors but not plant characteristics mainly alter soil microbial properties along a restoration chronosequence of Pinus tabulaeformis stands on Mt. Ziwuling, China. For. Ecol. Manag. 453, 117625. .
[54]
Zhou, L.L., Cai, L.P., He, Z.M., Wang, R.W., Wu, P.F., Ma, X.Q., 2016. Thinning increases understory diversity and biomass, and improves soil properties without decreasing growth of Chinese fir in southern China. Environ. Sci. Pollut. Res. 23, 24135-24150. .
Table
1.
Data classification structure of species richness of understory plants and regeneration density.
Thinning intensity (%)
Recovery time (year)
Age group (year)
HI
Thinning frequency
Species richness of understory plants
WT (≤30)
1–3
Young (≤20)
<30
Single thinning
MT (30–55)
4–6
Half-mature (21–30)
≥30
Multiple thinning
ST (>55)
7–13
Near-mature (31–40)
≥14
Mature (41–60)
Regeneration density
WT (≤30)
1–5
Half-mature (21–30)
<30
Single thinning
MT (30–50)
6–10
Near-mature (31–40)
≥30
Multiple thinning
ST (>50)
≥11
Mature (41–60)
Note: WT, MT and ST represented weak thinning, moderate thinning and strong thinning, respectively. The age groups were identified by “Regulations for age-class and age-group division of the main tree-species” (Regulatory document from the Chinese government, 2017, ICS 65.020 B60 LY, LY/T 2908-2017).
Table
2.
Factors affecting the species richness of understory plants in Chinese pine plantation.
Factors
%IncMSE
Total
Shrub
Herb
MAT
6.64
3.20
6.64
MAP
3.31
3.26
3.25
Aspect
2.52
1.49
3.49
Slope
4.86
8.59
6.36
Elevation
8.93
6.07
10.83
Planting density
11.42
11.85
9.85
Retention density
3.01
0.58
7.24
Age group
11.68
3.98
14.63
Thinning frequency
4.43
3.44
7.33
Thinning intensity
−1.00
−2.76
−0.10
Recovery time
9.88
4.31
9.68
Sample area
3.63
0.00
Note: MAT = mean annual temperature; MAP = mean annual precipitation. These statistics were evaluated by the randomForest function in randomForest package of R.
Table
3.
Classification statistics on the correlations between thinning and the species richness of understory plants and natural regeneration density.
Variable
Sample size (N)
+
–
0
Species richness of understory plants
RR-T
83
75 (90.36%)
7 (8.43%)
1 (1.20%)
RR-S
83
62 (74.70%)
12 (14.46%)
9 (10.84%)
RR-H
81
71 (87.65%)
9 (11.11%)
1 (1.23%)
Regeneration density
RR-d
43
33 (76.74%)
10 (23.26%)
0
Note: The symbols +, −, and 0 represent positive relationships, negative relationships, and no relationship, respectively. N represents the number of RR. RR-T, RR-S, RR-H represent the response ratio of the species richness of all understory plants, shrubs and herbs. RR-d represents the response ratio of regeneration density.

 DownLoad:
DownLoad:





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds