| [1] |
Adler, P.B., Smull, D., Beard, K.H., Choi, R.T., Furniss, T., Kulmatiski, A., Meiners, J.M., Tredennick, A.T., Veblen, K.E., 2018. Competition and coexistence in plant communities: intraspecific competition is stronger than interspecific competition. Ecol. Lett. 21, 1319-1329. .
|
| [2] |
Bai, X.J., Queenborough, S.A., Wang, X.G., Zhang, J., Li, B.H., Yuan, Z.Q., Xing, D.L., Lin, F., Ye, J., Hao, Z.Q., 2012. Effects of local biotic neighbors and habitat heterogeneity on tree and shrub seedling survival in an old-growth temperate forest. Oecologia 170, 755-765. .
|
| [3] |
Bates, D., Maechler, M., Bolker, B., Walker, S., 2015. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1-48. .
|
| [4] |
Bennett, J.A., Maherali, H., Reinhart, K.O., Lekberg, Y., Hart, M.M., Klironomos, J., 2017. Plant-soil feedbacks and mycorrhizal type influence temperate forest population dynamics. Science 355, 181-184. .
|
| [5] |
Barry, K.E., Schnitzer, S.A., 2021. Are we missing the forest for the trees? Conspecific negative density dependence in a temperate deciduous forest. PLoS One 16(7), e0245639. .
|
| [6] |
Bolker, B.M., Brooks, M.E., Clark, C.J., Geange, S.W., Poulsen, J.R., Stevens, M.H., White, J.S., 2009. Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol. Evol. 24, 127-135. .
|
| [7] |
Brown, A.J., Payne, C.J., White, P.S., Peet, R.K., 2020. Shade tolerance and mycorrhizal type may influence sapling susceptibility to conspecific negative density dependence. J. Ecol. 108, 325-336. .
|
| [8] |
Chen, L., Swenson, N.G., Ji, N.N., Mi, X.C., Ren, H.B., Guo, L.D., Ma, K.P., 2019. Differential soil fungus accumulation and density dependence of trees in a subtropical forest. Science 366 (6461), 124-128. .
|
| [9] |
Chi, X.L., Tang, Z.Y., Xie, Z.Q., Guo, Q., Zhang, M., Ge, J.L., Xiong, G.M., Fang, J.Y., 2015. Effects of size, neighbors, and site condition on tree growth in a subtropical evergreen and deciduous broad-leaved mixed forest, China. Ecol. Evol. 5, 5149-5161. .
|
| [10] |
Comita, L.S., Muller-Landau, H.C., Aguilar, S., Hubbell, S.P., 2010. Asymmetric density dependence shapes species abundances in a tropical tree community. Science 329, 330-332. .
|
| [11] |
Comita, L.S., Queenborough, S.A., Murphy, S.J., Eck, J.L., Xu, K., Krishnadas, M., Zhu, Y., 2014. Testing predictions of the Janzen-Connell hypothesis: a meta-analysis of experimental evidence for distance- and density-dependent seed and seedling survival. J. Ecol. 102, 845-856. .
|
| [12] |
Comita, L.S., 2017. How latitude affects biotic interactions. Science 356, 1328-1329. .
|
| [13] |
Comita, L.S., Hubbell, S.P., 2009. Local neighborhood and species' shade tolerance influence survival in a diverse seedling bank. Ecology 90, 328-334. .
|
| [14] |
Comita, L.S., Uriarte, M., Thompson, J., Jonckheere, I., Canham, C.D., Zimmerman, J.K., 2009. Abiotic and biotic drivers of seedling survival in a hurricane-impacted tropical forest. J. Ecol. 97, 1346-1359. .
|
| [15] |
Connell, J.H., 1971. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees, in: den Boer, P. J., Gradwell, O.R. (Eds. ), Dynamics of populations. Center for Agricultural Publishing and Documentation, Wageningen, pp. 298-312.
|
| [16] |
Crawford, K.M., Bauer, J.T., Comita, L.S., Eppinga, M.B., Johnson, D.J., Mangan, S.A., Queenborough, S.A., Strand, A.E., Suding, K.N., Umbanhowar, J., Bever, J.D., 2019. When and where plant-soil feedback may promote plant coexistence: a meta-analysis. Ecol. Lett. 22, 1274-1284. .
|
| [17] |
Fan, C.Y., Tan, L.Z., Zhang, P., Liang, J.J., Zhang, C.Y., Wang, J., Zhao, X.H., von Gadow, K., 2017. Determinants of mortality in a mixed broad-leaved Korean pine forest in northeastern China. Eur. J. Forest. Res. 136(3), 457-469. .
|
| [18] |
Fien, E.K.P., Fraver, S., Teets, A., Weiskittel, A.R., Hollinger, D.Y., 2019. Drivers of individual tree growth and mortality in an uneven-aged, mixed-species conifer forest. Forest Ecol. Manag. 449, 117446. .
|
| [19] |
Fine, P.V.A., Miller, Z.J., Mesones, I., Irazuzta, S., Appel, H.M., Stevens, M.H.H., Saaksjarvi, I., Schultz, J.C., Coley, P.D., 2006. The growth-defense trade-off and habitat specialization by plants in Amazonian forests. Ecology 87, S150-S162. .
|
| [20] |
Gadow, K.v., Kotze, H., 2014. Tree survival and maximum density of planted forests - Observations from South African spacing studies. For. Ecosyst. 1, 21. .
|
| [21] |
Iida, Y., Poorter, L., Sterck, F., Kassim, A.R., Potts, M.D., Kubo, T., Kohyama, T.S., 2014. Linking size-dependent growth and mortality with architectural traits across 145 co-occurring tropical tree species. Ecology 95, 353-363. .
|
| [22] |
Janzen, D.H., 1970. Herbivores and the number of tree species in tropical forests. Am. Nat. 104, 501-528. .
|
| [23] |
Jia, S.H., Wang, X.G., Yuan, Z.Q., Lin, F., Ye, J., Lin, G.G., Hao, Z.Q., Bagchi, R., 2020. Tree species traits affect which natural enemies drive the Janzen-Connell effect in a temperate forest. Nat. Commun. 11, 286. .
|
| [24] |
Jiang, F., Zhu, K., Cadotte, M.W., Jin, G.Z., 2020. Tree mycorrhizal type mediates the strength of negative density dependence in temperate forests. J. Ecol. 108, 2601-2610. .
|
| [25] |
Johnson, D.J., Beaulieu, W.T., Bever, J.D., Clay, K., 2012. Conspecific negative density dependence and forest diversity. Science 336, 904-907. .
|
| [26] |
Keram, A., Halik, U., Aishan, T., Keyimu M., Jiapaer K., Li G., 2021. Tree mortality and regeneration of Euphrates poplar riparian forests along the Tarim River, Northwest China. For. Ecosyst. 8, 49. .
|
| [27] |
Krueger, L.M., Peterson, C.J., Royo, A., Carson, W.P., 2009. Evaluating relationships among tree growth rate, shade tolerance, and browse tolerance following disturbance in an eastern deciduous forest. Can. J. Forest Res. 39(12), 2460-2469. .
|
| [28] |
Kuang, X., Zhu, K., Yuan, Z.Q., Lin, F., Ye, J., Wang, X.G., Wang, Y.Y., Hao, Z.Q., 2017. Conspecific density dependence and community structure: insights from 11 years of monitoring in an old-growth temperate forest in Northeast China. Ecol. Evol. 7 (14), 5191-5200. .
|
| [29] |
LaManna, J.A., Mangan, S.A., Alonso, A., Bourg, N.A., Brockelman, W.Y., Bunyavejchewin, S., Chang, L., Chiang, J.M., Chuyong, G.B., Clay, K., Condit, R., Cordell, S., Davies, S.J., Furniss, T.J., Giardina, C.P., Gunatilleke, I., Gunatilleke, C.V.S., He, F., Howe, R.W., Hubbell, S.P., Hsieh, C.F., Inman-Narahari, F.M., Janik, D., Johnson, D.J., Kenfack, D., Korte, L., Kral, K., Larson, A.J., Lutz, J.A., McMahon, S.M., McShea, W.J., Memiaghe, H.R., Nathalang, A., Novotny, V., Ong, P.S., Orwig, D.A., Ostertag, R., Parker, G.G., Phillips, R.P., Sack, L., Sun, I.F., Tello, J.S., Thomas, D.W., Turner, B.L., Vela Diaz, D.M., Vrska, T., Weiblen, G.D., Wolf, A., Yap, S., Myers, J.A., 2017. Plant diversity increases with the strength of negative density dependence at the global scale. Science 356, 1389-1392. .
|
| [30] |
Liu, X., Burslem, D.F.R.P., Taylor, J.D., Taylor, A.F.S., Khoo, E., Majalap-Lee, N., Helgason, T., Johnson, D., 2018. Partitioning of soil phosphorus among arbuscular and ectomycorrhizal trees in tropical and subtropical forests. Ecol. Lett. 21, 713-723. .
|
| [31] |
Liu, X.B., Etienne, R.S., Liang, M.X., Wang, Y., Yu, S.X., 2015. Experimental evidence for an intraspecific Janzen-Connell effect mediated by soil biota. Ecology 96, 662-671. .
|
| [32] |
Magee, L., Wolf, A., Howe, R., Schubbe, J., Hagenow, K., Turner, B., 2021. Density dependence and habitat heterogeneity regulate seedling survival in a North American temperate forest, Forest Ecol. Manag. 480, 118722. .
|
| [33] |
Mao, Z.K., Corrales, A., Zhu, K., Yuan, Z.Q., Lin, F., Hao, Z.Q., Wang, X.G., 2019. Tree mycorrhizal associations mediate soil fertility effects on forest community structure in a temperate forest. New Phytol. 223, 475-486. .
|
| [34] |
Niinemets, U., Valladares, F., 2006. Tolerance to shade, drought, and waterlogging of temperate Northern Hemisphere trees and shrubs. Ecol. Monogr. 76, 521-547. .
|
| [35] |
Piao, T.F., Comita, L.S., Jin, G.Z., Kim, J.H., 2013. Density dependence across multiple life stages in a temperate old growth forest of northeast China. Oecologia 172(1), 207-217. .
|
| [36] |
Qin, J.H., Geng, Y., Li, X.Y., Zhang, C.Y., Zhao, X.H., von Gadow, K., 2021. Mycorrhizal type and soil pathogenic fungi mediate tree survival and density dependence in a temperate forest. Forest Ecol. Manag. 496, 119459. .
|
| [37] |
Qin, J.H., Zhang, Z.H., Geng, Y., Zhang, C.Y., Song, Z.L., Zhao, X.H., 2020. Variations of density-dependent seedling survival in a temperate forest. Forest Ecol. Manag. 468, 118158. .
|
| [38] |
Queenborough, S.A., Metz, M.R., Valencia, R., Wright, S.J., 2013. Demographic consequences of chromatic leaf defence in tropical tree communities: do red young leaves increase growth and survival? Ann. Bot-London. 112(4), 677-684. .
|
| [39] |
R Core Team, 2021. R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria.
|
| [40] |
Ramage, B.S., Mangana, I.J., 2017. Conspecific negative density dependence in American beech. For. Ecosyst. 4, 8. .
|
| [41] |
Rees, M., Condit, R., Crawley, M., Pacala, S., Tilman, D., 2001. Long-term studies of vegetation dynamics. Science 293, 650-655. .
|
| [42] |
Ruger, N., Wirth, C., Wright, S.J., Condit, R., 2012. Functional traits explain light and size response of growth rates in tropical tree species. Ecology 93, 2626-2636. .
|
| [43] |
Russo, S.E., Wiser, S.K., Coomes, D.A., 2007. Growth-size scaling relationships of woody plant species differ from predictions of the Metabolic Ecology Model. Ecol. Lett. 10(10), 889-901. .
|
| [44] |
Russo, S.E., Jenkins, K.L., Wiser, S.K., Uriarte, M., Duncan, R.P., Coomes, D.A., 2010. Interspecific relationships among growth, mortality and xylem traits of woody species from New Zealand. Funct. Ecol. 24(2), 253-262. .
|
| [45] |
Russo, S.E., McMahon, S.M., Detto, M., Ledder, G., Wright, S.J., Condit, R.S., Davies, S.J., Ashton, P.S., Bunyavejchewin, S., Chang-Yang, C-H., Ediriweera, S., Ewango, C.E.N., Fletcher, C., Foster, R.B., Gunatilleke, C.V.S., Gunatilleke, I.A.U.N., Hart, T., Hsieh, C-F., Hubbell, S.P., Itoh, A., Kassim, A.R., Leong, Y.T., Lin, Y.C., Makana, J-R., Mohamad, M.Bt., Ong, P., Sugiyama, A., Sun, I-F., Tan, S., Thompson, J., Yamakura, T., Yap, S.L., Zimmerman, J.K., 2021. The interspecific growth-mortality trade-off is not a general framework for tropical forest community structure. Nat. Ecol. Evol. 5, 174-183. .
|
| [46] |
Song, X.Y., Lim, J.Y., Yang, J., Luskin, M.S., 2021. When do Janzen-Connell effects matter? A phylogenetic meta-analysis of conspecific negative distance and density dependence experiments. Ecol. Lett. 24, 608-620. .
|
| [47] |
Valladares, F., Niinemets, Ü., 2008. Shade tolerance, a key plant feature of complex nature and consequences. Ann. Rev. Ecol. Evol. S39, 237-257. .
|
| [48] |
Wang, X.G., Comita, L.S., Hao, Z.Q., Davies, S.J., Ye, J., Lin, F., Yuan, Z.Q., 2012. Local-scale drivers of tree survival in a temperate forest. PLoS One 7, e29469. .
|
| [49] |
Wang, X.G., Hao, Z.Q., Zhang, J., Lian, J.Y., Li, B.H., Ye, J., Yao, X.L., 2009. Tree size distributions in an old-growth temperate forest. Oikos 118, 25-36. .
|
| [50] |
Wright, S.J., Kitajima, K., Kraft, N.J.B., Reich, P.B., Wright, I.J., Bunker, D.E., Condit, R., Dalling, J.W., Davies, S.J., Diaz, S., Engelbrecht, B.M.J., Harms, K.E., Hubbell, S.P., Marks, C.O., Ruiz-Jaen, M.C., Salvador, C.M., Zanne, A.E., 2010. Functional traits and the growth-mortality trade-off in tropical trees. Ecology 91, 3664-3674. .
|
| [51] |
Wu, H., Franklin, S.B., Liu, J., Lu, Z., 2017. Relative importance of density dependence and topography on tree mortality in a subtropical mountain forest. Forest Ecol. Manag. 384, 169-179. .
|
| [52] |
Wunder, J., Reineking, B., Bigler, C., Bugmann, H., 2008. Predicting tree mortality from growth data: how virtual ecologists can help real ecologists. J. Ecol. 96, 174-187. .
|
| [53] |
Yan, Y., Zhang, C.Y., Wang, Y.X., Zhao, X.H., von Gadow, K., 2015. Drivers of seedling survival in a temperate forest and their relative importance at three stages of succession. Ecol. Evol. 5, 4287-4299. .
|
| [54] |
Yao, J., Bachelot, B., Meng, L.J., Qin, J.H., Zhao, X.H., Zhang, C.Y., 2020. Abiotic niche partitioning and negative density dependence across multiple life stages in a temperate forest in northeastern China. J. Ecol. 108, 1299-1310. .
|
| [55] |
Zhu, K., Woodall, C.W., Monteiro, J.V.D., Clark, J.S., 2015a. Prevalence and strength of density-dependent tree recruitment. Ecology 96, 2319-2327. .
|
| [56] |
Zhu, Y., Comita, L.S., Hubbell, S.P., Ma, K.P., 2015b. Conspecific and phylogenetic density-dependent survival differs across life stages in a tropical forest. J. Ecol. 103, 957-966. .
|
| [57] |
Zhu, Y., Queenborough, S.A., Condit, R., Hubbell, S.P., Ma, K.P., Comita, L.S., 2018. Density-dependent survival varies with species life-history strategy in a tropical forest. Ecol. Lett. 21, 506-515. .
|
| [58] |
Zhu, Y., Hogan, J.A., Cai, H.Y., Xun, Y.H., Jiang, F., Jin, G.Z., 2017. Biotic and abiotic drivers of the tree growth and mortality trade-off in an old-growth temperate forest. Forest Ecol. Manag. 404, 354-360. .
|

 DownLoad:
DownLoad:
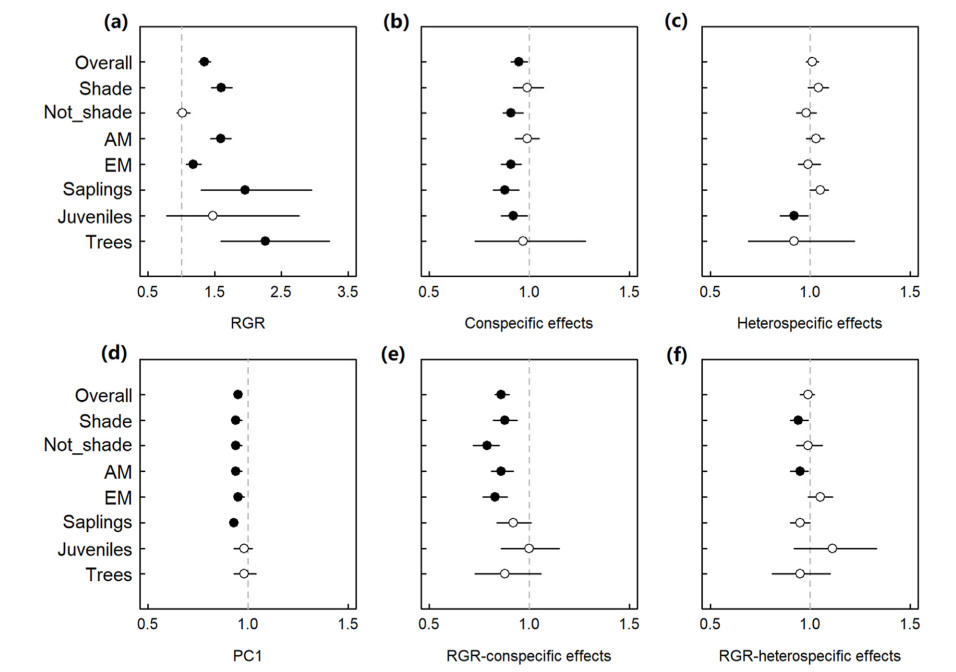
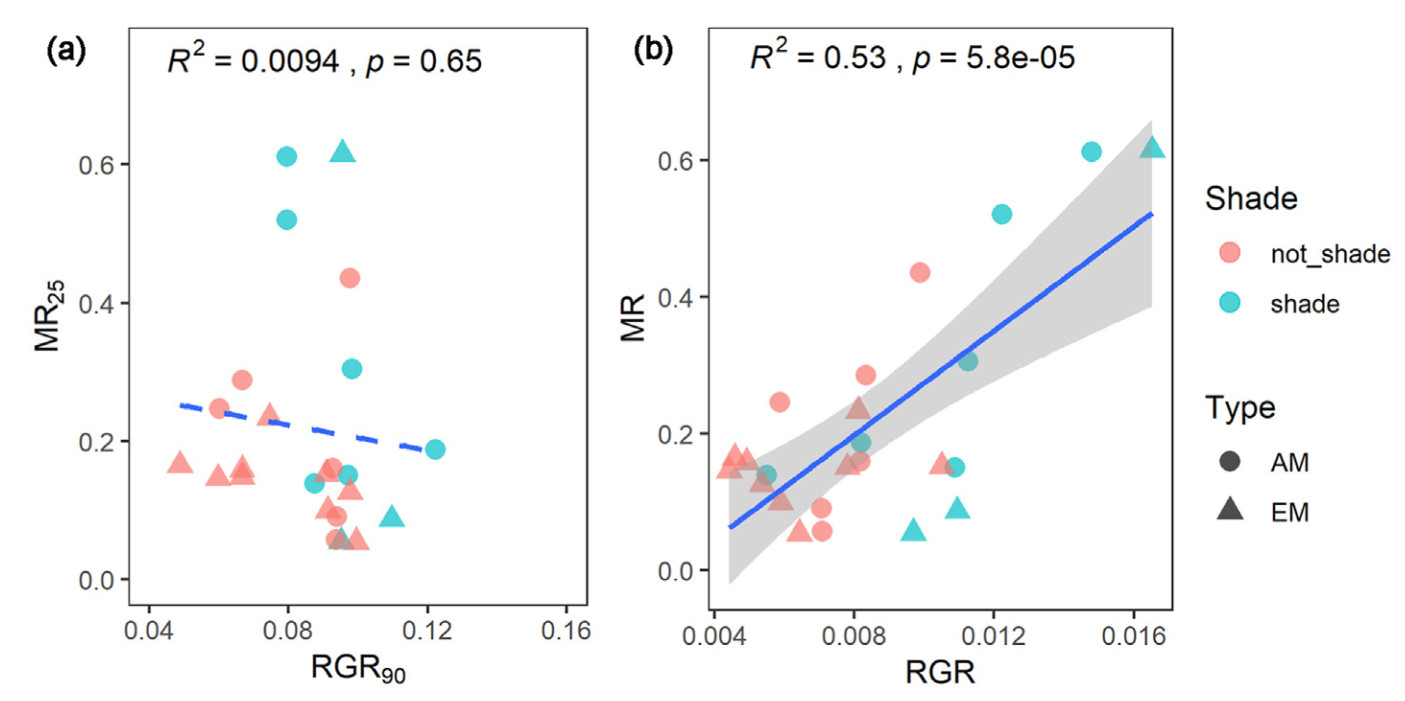
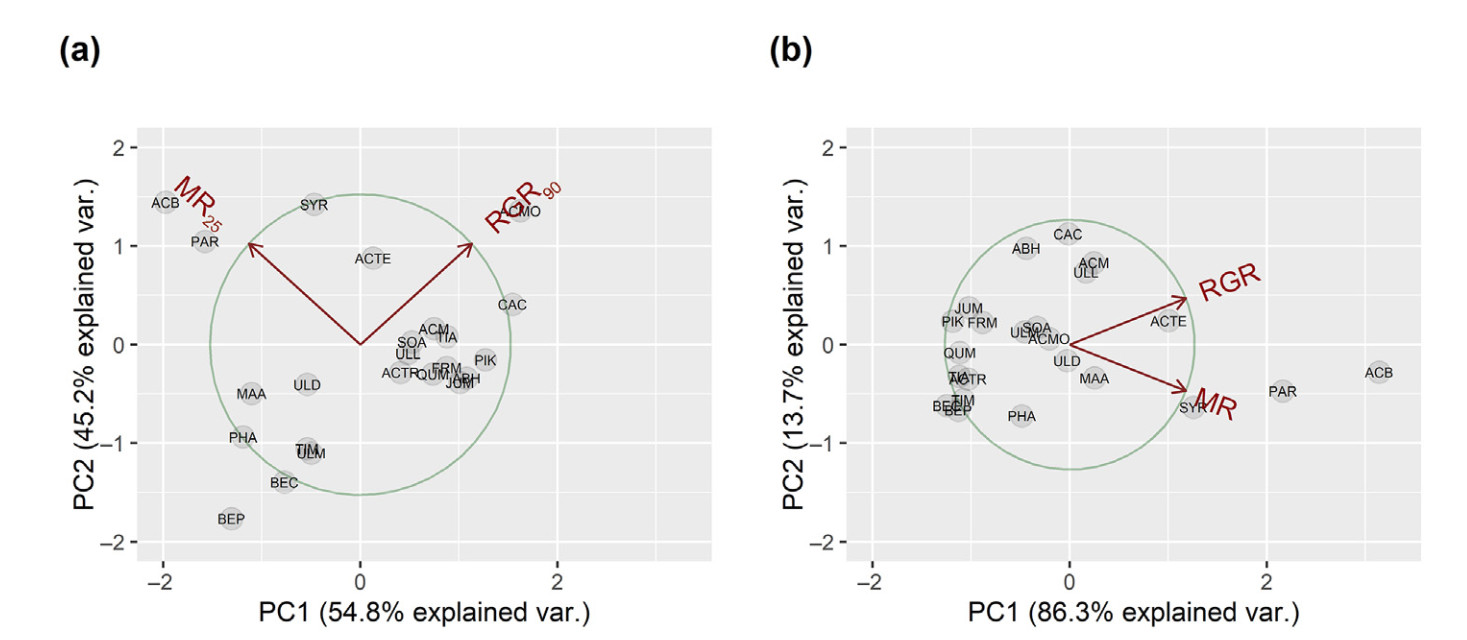
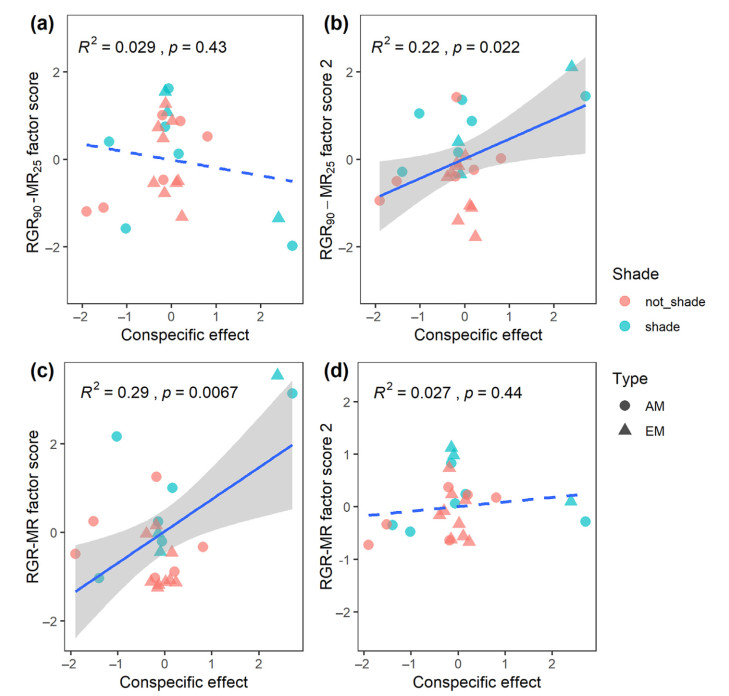





 Email Alerts
Email Alerts RSS Feeds
RSS Feeds